Файл: Иммунная система человека представлена комплексом лимфомиелоидных органов и лимфоидной ткани, ассоциированной с дыхательной, пищеварительной и мочеполовой системами.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 07.11.2023
Просмотров: 338
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
СОДЕРЖАНИЕ
Миграция Т-лимфоцитов на периферию
Развитие В-лимфоцитов в костном мозге
Строение В-клеточного рецептора
Развитие мононуклеарных фагоцитов в костном мозге и тканях
Эозинофильные и базофильные гранулоциты
Морфология и клеточный состав лимфоидных образований слизистых
Закономерности развития иммунных реакций в слизистых оболочках
Биологическая характеристика цитокинов
Роль системы комплемента при болезнях
ЦИК – циркулирующие иммунные комплексы
ГЛАВНЫЙ КОМПЛЕКС ГИСТОСОВМЕСТИМОСТИ: СТРУКТУРА И ФУНКЦИИ
Эндотелиальные клетки. Миграция лейкоцитов в зону воспаления
Иммунологические методы исследования
+-B-лимфоцитов. Фолликулярная зона включает зародышевый центр, в котором происходит пролиферация В-клеток и их дифференцировка в плазматические клетки – продуценты IgA. Т-клеточная зона содержит две основные Т-клеточные субпопуляции: Т-хелперы и Т-цитотоксичекие лимфоциты.
Функционально в иммунной системе слизистых покровов выделяют два участка: индуктивный (лимфатические узлы, миндалины, лимфоидные бляшки, лимфоидные узелки) и эффекторный (lamina propria и межэпителиальные лимфоциты). В индуктивном участке происходит распознавание антигена, его презентация в иммуногенной форме и формирование популяции антигенспецифических лимфоцитов. В этом процессе принимают участие дендритные клетки, макрофаги, ГКГ II+-клетки, CD4+, -лимфoциты, СD8+-лимфоциты, sIgA+-B-лимфоциты.
В эффекторном участке продуцируется секреторный IgA и накапливаются эффекторные Т-лимфоциты, которые обеспечивают клеточную защиту слизистых поверхностей. В lamina propria от общего количества лимфоидных клеток Т-лимфоциты составляют 40-60%, В-лимфоциты – 20-40%, НК-клетки – 2-3%; межэпителиальные лимфоциты на 80-90% представлены Т-клетками, среди которых основной процент принадлежит СD8+-лимфоцитам, обладающих цитотоксичностью и способностью усиливать продукцию sIgA.
Проникновение патогена в слизистую оболочку приводит к его захвату макрофагами и презентации Т- и В-лимфоцитам. Из просвета кишечника антигены доставляются в индуктивный отдел иммунной системы слизистой с помощью М-клеток, где в дальнейшем представляются иммунокомпетентным клеткам.
Под влиянием антигена происходит отбор клоноспецифических Т- и В-лимфоцитов , их активация и пролиферация. Большая часть активированных Т- и В-лимфоцитов проходит своё развитие до зрелых эффекторных клеток (плазматических или цитотоксических клеток) в индуктивном отделе иммунной системы слизистых (лимфоидных узелках, лимфоидных бляшках, регионарных лимфатических узлах), после чего эти клетки мигрируют в эффекторную зону иммунной системы слизистых, где и реализуют свои свойства (продуцируют IgA или вызывают цитолиз чужеродных клеток).
Основную роль в регуляции дифференцировки IgA+ В-клеток в антитело-секретирующие клетки играют ТН2-хелперы (CD4+, ), и продуцируемые ими ИЛ-5, ИЛ-6.
Часть антигенактивированных в слизистых покровах Т- и В-лимфоцитов через лимфатические сосуды и грудной лимфатический проток попадает в системную циркуляцию, откуда они диссеминируют в селезенку, отдаленные лимфатические узлы и индуктивные участки лимфоидных образований слизистых других систем организма. Такая высокая миграционная активность лимфоидных клеток слизистых обеспечивает непрерывное пополнение всех участков слизистых покровов (желудочно-кишечного тракта, респираторной системы, мочеполовой системы и железистых органов) антигенреактивными Т- и В-лимфоцитами, что в итоге служит постоянному поддержанию высокой общей иммунореактивности организма.
Особую роль в этом процессе играет иммунная система желудочно-кишечного тракта, которая, как известно, находится в постоянном контакте с громадным потоком микробного и аллергенного материала. Установлено, что пейеровы бляшки тонкой кишки являются важным (но не единственным) источником плазмоцитов, секретирующих IgA, практически для всех слизистых оболочек и железистых органов, а стимуляция иммунокомпетентных клеток пейеровых бляшек ведет к активации иммунной системы не только ЖКТ, но и дыхательного и урогенитального трактов. Известно, что секреторный IgA обеспечивает эффективную защиту слизистых покровов от инвазии микробов, вирусов, от проникновения токсинов и аллергенов, усиливает антибактериальную активность фагоцитов. Селективный дефицит IgA, развивающийся по разным причинам, в том числе и вследствие мутации в структурных генах Нa-цепи, способен приводить к аллергическим, инфекционным и аутоиммунным заболеваниям кишечника и других слизистых структур организма. Установлено, что повышенный поток микробных антигенов и токсинов может вести к активации «дремлющих» клонов аутоиммунных лимфоцитов и при наличии соответствующих условий может служить пусковым механизмом в развитии патологического процесса. Отсюда следует, что нормализация работы иммунной системы слизистых покровов и поддержание нормальной продукции IgA является обязательным условием лечения и профилактики инфекционных, аллергических и аутоиммунных заболеваний.
Антитела — специфические белки гамма- глобулиновой природы, образующиеся в организме в ответ на антигенную стимуляцию и способные специфически взаимодействовать с антигеном (in vivo, in vitro). В соответствии с международной классификацией совокупность сывороточных белков, обладающих свойствами антител, называют иммуноглобулинами.
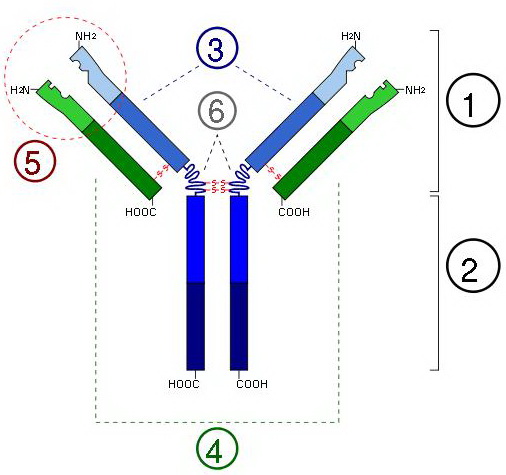
Общий план строения иммуноглобулинов:
1) Fab (Участок связывания антигена (Fab, fragment antigen binding) — это участок молекулы иммуноглобулина, который связывает антиген. Fab состоит из одного константного и одного вариабельного домена легкой и тяжелой цепей. Эти домены образуют паратоп — сайт связывания антигена на N-конце. Вариабельные домены связывают эпитоп специфических антигенов). Fc и Fab могут быть получены разрезанием молекулы иммуноглобулина ферментом папаином, при этом из одной молекулы мономерного иммуноглобулина образуются два Fab и один Fc. Фермент пепсин разрезает иммуноглобулины ниже шарнирного участка и при этом образуется один мономер Fc и один F(ab)2);
2) Fc (Кристаллизующийся фрагмент иммуноглобулина (Fc, fragment crystallizable region, Fc region) — это концевая часть молекулы иммуноглобулина, которая взаимодействует с Fc рецептором на поверхности клетки и с некоторыми белками системы комплемента. Данное свойство позволяет антителам активировать иммунную систему. Fc участок IgG, IgA и IgD изотипов состоит из двух одинаковых белковых фрагментов, соответственно, второго и третьего константных доменов двух тяжелых цепей; в случае изотипов IgM и IgE Fc содержит три константных домена тяжелых цепей (домены CH 2-4) в каждой полипептидной цепочке. Другая часть антитела называется Fab (от англ. Fragment antigen binding), и состоит из вариабельных участков, которые определяют специфичность мишени, которую связывает антитело. Напротив, Fc всех антител одного класса одинаковы и константны. Отсюда происходит
неправильное объяснение происхождения термина Fc как «fragment constant region»);
3) тяжелая цепь;
4) легкая цепь;
5) антиген-связывающийся участок;
6) шарнирный участок.
Уникальность антител заключается в том, что они способны специфически взаимодействовать только с тем антигеном, который вызвал их образование.
Иммуноглобулины ( Ig ) разделены в зависимости от локализации на три группы:
- сывороточные (в крови);
- секреторные ( в секретах- содержимом желудочно- кишечного тракта, слезном секрете, слюне, особенно- в грудном молоке) обеспечивают местный иммунитет (иммунитет слизистых);
- поверхностные ( на поверхности иммунокомпетентных клеток, особенно В- лимфоцитов).
Любая молекула антител имеет сходное строение ( Y- образную форму) и состоит из двух тяжелых ( Н ) и двух легких ( L ) цепей, связанных дисульфидными мостиками. Каждая молекула антител имеет два одинаковых антигенсвязывающих фрагмента Fab ( fragment antigen binding ), определяющих антительную специфичность, и один Fc ( fragment constant ) фрагмент, который не связывает антиген, но обладает эффекторными биологическими функциями. Он взаимодействует со “своим” рецептором в мембране различных типов клеток ( макрофаг, тучная клетка, нейтрофил).
Концевые участки легких и тяжелых цепей молекулы иммуноглобулина вариабельны по составу ( аминокислотным последовательностям ) и обозначаются как VL и VH области. В их составе выделяют гипервариабельные участки, которые определяют структуру активного центра антител (антигенсвязывающий центр или паратоп). Именно с ним взаимодействует антигенная детерминанта (эпитоп) антигена. Антигенсвязывающий центр антител комплементарен эпитопу антигена по принципу “ключ — замок” и образован гипервариабельными областями L- и Н- цепей. Антитело свяжется антигеном (ключ попадет в замок) только в том случае, если детерминантная группа антигена полностью вместится в щель активного центра антител.
Легкие и тяжелые цепи состоят из отдельных блоков- доменов
. В легких ( L ) цепях — два домена- один вариабельный ( V ) и один константный ( C ), в тяжелых ( H ) цепях- один V и 3 или 4 ( в зависимости от класса иммуноглобулина ) C домена.
Существуют легкие цепи двух типов- каппа и лямбда, они встречаются в различных пропорциях в составе различных (всех) классов иммуноглобулинов.
Выявлено пять классов тяжелых цепей- альфа ( с двумя подклассами), гамма ( с четырьмя подклассами), эксилон, мю и дельта. Соответственно обозначению тяжелой цепи обозначается и класс молекул иммуноглобулинов- А, G, E, M и D.
Именно константные области тяжелых цепей, различаясь по аминокислотному составу у различных классов иммуноглобулинов, в конечном результате и определяют специфические свойства иммуноглобулинов каждого класса.
Известно пять классов иммуноглобулинов, отличающихся по строению тяжелых цепей, молекулярной массе, физико- химическим и биологическим характеристикам: IgG, IgM, IgA, IgE, IgD. В составе IgG выделяют 4 подкласса ( IgG1, IgG2, IgG3, IgG4 ), в составе IgA- два подкласса (IgA1, IgA2 ).
Структурной единицей антител является мономер, состоящий из двух легких и двух тяжелых цепей. Мономерами являются IgG, IgA ( сывороточный), IgD и IgE. IgM- пентамер (полимерный Ig). У полимерных иммуноглобулинов имеется дополнительная j ( joint) полипептидная цепь, которая объединяет ( полимеризует) отдельные субъединицы (в составе пентамера IgM, ди- и тримера секреторного IgA).
Основные биологические характеристики антител.
1. Специфичность — способность взаимодействия с определенным (своим) антигеном (соответствие эпитопа антигена и активного центра антител).
2. Валентность- количество способных реагировать с антигеном активных центров ( это связано с молекулярной организацией- моно- или полимер). Иммуноглобулины могут быть двухвалентными ( IgG ) или поливалентными (пентамер IgM имеет 10 активных центров). Двух- и более валентные антитела навывают полными антителами. Неполные антитела
Функционально в иммунной системе слизистых покровов выделяют два участка: индуктивный (лимфатические узлы, миндалины, лимфоидные бляшки, лимфоидные узелки) и эффекторный (lamina propria и межэпителиальные лимфоциты). В индуктивном участке происходит распознавание антигена, его презентация в иммуногенной форме и формирование популяции антигенспецифических лимфоцитов. В этом процессе принимают участие дендритные клетки, макрофаги, ГКГ II+-клетки, CD4+, -лимфoциты, СD8+-лимфоциты, sIgA+-B-лимфоциты.
В эффекторном участке продуцируется секреторный IgA и накапливаются эффекторные Т-лимфоциты, которые обеспечивают клеточную защиту слизистых поверхностей. В lamina propria от общего количества лимфоидных клеток Т-лимфоциты составляют 40-60%, В-лимфоциты – 20-40%, НК-клетки – 2-3%; межэпителиальные лимфоциты на 80-90% представлены Т-клетками, среди которых основной процент принадлежит СD8+-лимфоцитам, обладающих цитотоксичностью и способностью усиливать продукцию sIgA.
11
Закономерности развития иммунных реакций в слизистых оболочках
Проникновение патогена в слизистую оболочку приводит к его захвату макрофагами и презентации Т- и В-лимфоцитам. Из просвета кишечника антигены доставляются в индуктивный отдел иммунной системы слизистой с помощью М-клеток, где в дальнейшем представляются иммунокомпетентным клеткам.
Под влиянием антигена происходит отбор клоноспецифических Т- и В-лимфоцитов , их активация и пролиферация. Большая часть активированных Т- и В-лимфоцитов проходит своё развитие до зрелых эффекторных клеток (плазматических или цитотоксических клеток) в индуктивном отделе иммунной системы слизистых (лимфоидных узелках, лимфоидных бляшках, регионарных лимфатических узлах), после чего эти клетки мигрируют в эффекторную зону иммунной системы слизистых, где и реализуют свои свойства (продуцируют IgA или вызывают цитолиз чужеродных клеток).
Основную роль в регуляции дифференцировки IgA+ В-клеток в антитело-секретирующие клетки играют ТН2-хелперы (CD4+, ), и продуцируемые ими ИЛ-5, ИЛ-6.
Часть антигенактивированных в слизистых покровах Т- и В-лимфоцитов через лимфатические сосуды и грудной лимфатический проток попадает в системную циркуляцию, откуда они диссеминируют в селезенку, отдаленные лимфатические узлы и индуктивные участки лимфоидных образований слизистых других систем организма. Такая высокая миграционная активность лимфоидных клеток слизистых обеспечивает непрерывное пополнение всех участков слизистых покровов (желудочно-кишечного тракта, респираторной системы, мочеполовой системы и железистых органов) антигенреактивными Т- и В-лимфоцитами, что в итоге служит постоянному поддержанию высокой общей иммунореактивности организма.
Особую роль в этом процессе играет иммунная система желудочно-кишечного тракта, которая, как известно, находится в постоянном контакте с громадным потоком микробного и аллергенного материала. Установлено, что пейеровы бляшки тонкой кишки являются важным (но не единственным) источником плазмоцитов, секретирующих IgA, практически для всех слизистых оболочек и железистых органов, а стимуляция иммунокомпетентных клеток пейеровых бляшек ведет к активации иммунной системы не только ЖКТ, но и дыхательного и урогенитального трактов. Известно, что секреторный IgA обеспечивает эффективную защиту слизистых покровов от инвазии микробов, вирусов, от проникновения токсинов и аллергенов, усиливает антибактериальную активность фагоцитов. Селективный дефицит IgA, развивающийся по разным причинам, в том числе и вследствие мутации в структурных генах Нa-цепи, способен приводить к аллергическим, инфекционным и аутоиммунным заболеваниям кишечника и других слизистых структур организма. Установлено, что повышенный поток микробных антигенов и токсинов может вести к активации «дремлющих» клонов аутоиммунных лимфоцитов и при наличии соответствующих условий может служить пусковым механизмом в развитии патологического процесса. Отсюда следует, что нормализация работы иммунной системы слизистых покровов и поддержание нормальной продукции IgA является обязательным условием лечения и профилактики инфекционных, аллергических и аутоиммунных заболеваний.
12
Антитела.
Антитела — специфические белки гамма- глобулиновой природы, образующиеся в организме в ответ на антигенную стимуляцию и способные специфически взаимодействовать с антигеном (in vivo, in vitro). В соответствии с международной классификацией совокупность сывороточных белков, обладающих свойствами антител, называют иммуноглобулинами.
Общий план строения иммуноглобулинов:
1) Fab (Участок связывания антигена (Fab, fragment antigen binding) — это участок молекулы иммуноглобулина, который связывает антиген. Fab состоит из одного константного и одного вариабельного домена легкой и тяжелой цепей. Эти домены образуют паратоп — сайт связывания антигена на N-конце. Вариабельные домены связывают эпитоп специфических антигенов). Fc и Fab могут быть получены разрезанием молекулы иммуноглобулина ферментом папаином, при этом из одной молекулы мономерного иммуноглобулина образуются два Fab и один Fc. Фермент пепсин разрезает иммуноглобулины ниже шарнирного участка и при этом образуется один мономер Fc и один F(ab)2);
2) Fc (Кристаллизующийся фрагмент иммуноглобулина (Fc, fragment crystallizable region, Fc region) — это концевая часть молекулы иммуноглобулина, которая взаимодействует с Fc рецептором на поверхности клетки и с некоторыми белками системы комплемента. Данное свойство позволяет антителам активировать иммунную систему. Fc участок IgG, IgA и IgD изотипов состоит из двух одинаковых белковых фрагментов, соответственно, второго и третьего константных доменов двух тяжелых цепей; в случае изотипов IgM и IgE Fc содержит три константных домена тяжелых цепей (домены CH 2-4) в каждой полипептидной цепочке. Другая часть антитела называется Fab (от англ. Fragment antigen binding), и состоит из вариабельных участков, которые определяют специфичность мишени, которую связывает антитело. Напротив, Fc всех антител одного класса одинаковы и константны. Отсюда происходит
неправильное объяснение происхождения термина Fc как «fragment constant region»);
3) тяжелая цепь;
4) легкая цепь;
5) антиген-связывающийся участок;
6) шарнирный участок.
Уникальность антител заключается в том, что они способны специфически взаимодействовать только с тем антигеном, который вызвал их образование.
Иммуноглобулины ( Ig ) разделены в зависимости от локализации на три группы:
- сывороточные (в крови);
- секреторные ( в секретах- содержимом желудочно- кишечного тракта, слезном секрете, слюне, особенно- в грудном молоке) обеспечивают местный иммунитет (иммунитет слизистых);
- поверхностные ( на поверхности иммунокомпетентных клеток, особенно В- лимфоцитов).
Любая молекула антител имеет сходное строение ( Y- образную форму) и состоит из двух тяжелых ( Н ) и двух легких ( L ) цепей, связанных дисульфидными мостиками. Каждая молекула антител имеет два одинаковых антигенсвязывающих фрагмента Fab ( fragment antigen binding ), определяющих антительную специфичность, и один Fc ( fragment constant ) фрагмент, который не связывает антиген, но обладает эффекторными биологическими функциями. Он взаимодействует со “своим” рецептором в мембране различных типов клеток ( макрофаг, тучная клетка, нейтрофил).
Концевые участки легких и тяжелых цепей молекулы иммуноглобулина вариабельны по составу ( аминокислотным последовательностям ) и обозначаются как VL и VH области. В их составе выделяют гипервариабельные участки, которые определяют структуру активного центра антител (антигенсвязывающий центр или паратоп). Именно с ним взаимодействует антигенная детерминанта (эпитоп) антигена. Антигенсвязывающий центр антител комплементарен эпитопу антигена по принципу “ключ — замок” и образован гипервариабельными областями L- и Н- цепей. Антитело свяжется антигеном (ключ попадет в замок) только в том случае, если детерминантная группа антигена полностью вместится в щель активного центра антител.
Легкие и тяжелые цепи состоят из отдельных блоков- доменов
. В легких ( L ) цепях — два домена- один вариабельный ( V ) и один константный ( C ), в тяжелых ( H ) цепях- один V и 3 или 4 ( в зависимости от класса иммуноглобулина ) C домена.
Существуют легкие цепи двух типов- каппа и лямбда, они встречаются в различных пропорциях в составе различных (всех) классов иммуноглобулинов.
Выявлено пять классов тяжелых цепей- альфа ( с двумя подклассами), гамма ( с четырьмя подклассами), эксилон, мю и дельта. Соответственно обозначению тяжелой цепи обозначается и класс молекул иммуноглобулинов- А, G, E, M и D.
Именно константные области тяжелых цепей, различаясь по аминокислотному составу у различных классов иммуноглобулинов, в конечном результате и определяют специфические свойства иммуноглобулинов каждого класса.
Известно пять классов иммуноглобулинов, отличающихся по строению тяжелых цепей, молекулярной массе, физико- химическим и биологическим характеристикам: IgG, IgM, IgA, IgE, IgD. В составе IgG выделяют 4 подкласса ( IgG1, IgG2, IgG3, IgG4 ), в составе IgA- два подкласса (IgA1, IgA2 ).
Структурной единицей антител является мономер, состоящий из двух легких и двух тяжелых цепей. Мономерами являются IgG, IgA ( сывороточный), IgD и IgE. IgM- пентамер (полимерный Ig). У полимерных иммуноглобулинов имеется дополнительная j ( joint) полипептидная цепь, которая объединяет ( полимеризует) отдельные субъединицы (в составе пентамера IgM, ди- и тримера секреторного IgA).
Основные биологические характеристики антител.
1. Специфичность — способность взаимодействия с определенным (своим) антигеном (соответствие эпитопа антигена и активного центра антител).
2. Валентность- количество способных реагировать с антигеном активных центров ( это связано с молекулярной организацией- моно- или полимер). Иммуноглобулины могут быть двухвалентными ( IgG ) или поливалентными (пентамер IgM имеет 10 активных центров). Двух- и более валентные антитела навывают полными антителами. Неполные антитела