Файл: Геном совокупность наследственного материала, заключенного в клетке организма. Геном содержит биологическую информацию, необходимую для выполнения всех функций организма..docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 22.11.2023
Просмотров: 20
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
рис. 6.

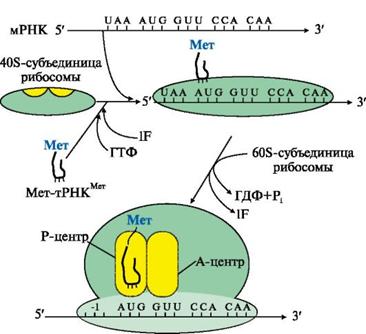
Рис. 6. Инициация белкового синтеза.
Этап элонгации включает три последовательные стадии:
1. связывание аа-тРНКаа в А-центре (требует затраты энергии ГТФ и участия фактора элонгации EF1 - на схеме аа-тРНКаа - Вал-тРНКВал);
2. образование пептидной связи катализирует пептидилтрансфераза, активный центр которой формируется рРНК, входящей в состав большой субъединицы рибосомы;
3. перемещение рибосомы по мРНК на один кодон в направлении от 5'- к 3'-концу идет с использованием энергии ГТФ (транслокация) и при участии фактора EF2, рис. 7.
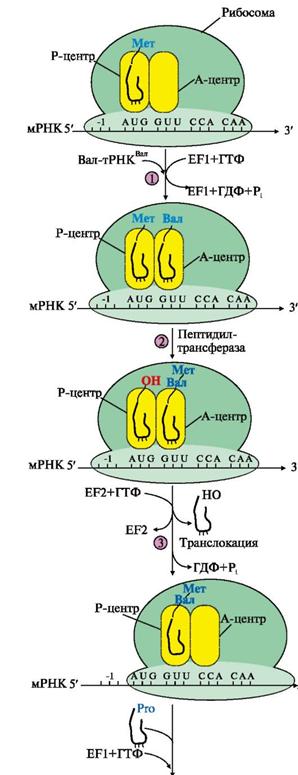
Рис. 7. Элонгация полипептидной цепи
Терминация трансляции происходит после включения в А-центр одного из стоп кодонов: UAG, UGA, UAA, рис. 8.
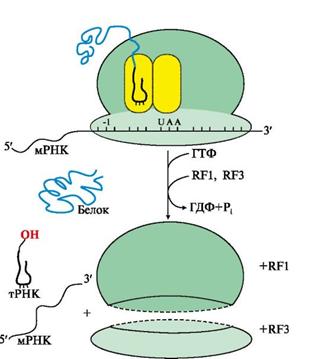
Рис. 8. Терминация синтеза белка
Одновременно несколько рибосом могут участвовать в трансляции одной мРНК. Каждая рибосома занимает участок, равный примерно 80 нуклеотидам мРНК. Таким образом, рибосомы располагаются на мРНК с интервалами около 100 нуклеотидов, образуя комплекс, называемый полисомой.
Адаптация организмов к различным воздействиям окружающей среды часто осуществляется путем изменения экспрессии (активности) генов, т.е. изменения скорости транскрипции определенных участков на молекулах ДНК. Этот процесс детально изучен на бактериях.
Адаптивная регуляция активности генов у прокариотов получила объяснение в теории оперона. Согласно этой теории на молекуле ДНК прокариотов присутствуют определенные участки - опероны.
В состав оперона входят:
1. структурные гены - содержат информацию о группе функционально взаимосвязанных белков, которые участвуют в одном и том же метаболическом пути;
2. промотор – расположен в начале оперона, короткая последовательность в несколько десятков нуклеотидов ДНК, с которой специфически связывается фермент РНК-полимераза
3. оператор - участок, с которым происходит связывание белка-репрессора.
4. терминатор – последовательность нуклеотидов, прекращающая транскрипцию.
Участки промотора и оператора частично перекрываются.
Белок-репрессор синтезируется в клетке с постоянной скоростью, его строение кодирует мРНК, транскрибируемая с гена-регулятора, расположенного на некотором расстоянии от оперона, работу которого контролирует его белковый продукт.
Если оперон регулируется по механизму индукции (например, лактозный оперон), то в отсутствии индуктора (лактозы) белок-репрессор связан с оператором. Комплекс белок-репрессор-оператор препятствует связыванию РНК-полимеразы с промотором, и транскрипция структурных генов оперона не идет (рис. 9 А).
Когда концентрация индуктора в клетке возрастет, то он присоединится к белку-репрессору, который имеет центр для связывания индуктора, изменяет его конформацию и снижает сродство к оператору. Комплекс индуктор-белок-репрессор теряет сродство к ДНК и уходит в цитозоль клетки. РНК-полимераза связывается с промотором и транскрибирует структурные гены. Идет синтез белков, закодированных в данном опероне (рис. 9 Б).
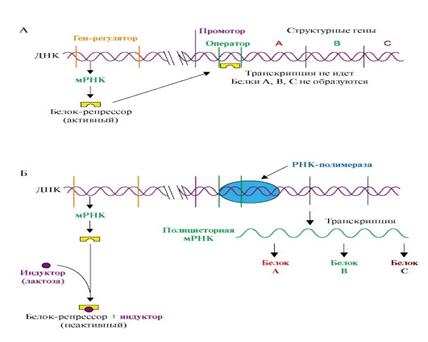
Рис. 9. Оперон, регулируемый по механизму индукции (лактозный оперон)
При регуляции оперона по механизму репрессии (например, гистидиновый или триптофановый опероны) белок-репрессор, постоянно синтезируемый в клетках, не имеет сродства к оператору (рис. 10 А). Когда к белку-репрессору присоединится небольшая молекула - корепрессор (гистидин или триптофан, как правило, конечный продукт метаболического пути, ферменты которого закодированы в структурных генах оперона), то в результате конформационных изменений комплекс белка-репрессора с корепрессором приобретает сродство к оператору и прекращает транскрипцию (рис. 10 Б). Так, гистидиновый оперон содержит 10 структурных генов, кодирующих строение ферментов синтеза гистидина. Добавки гистидина в среду выращивания клеток E. Coli вызывают снижение количества, а затем и полное исчезновение из внутриклеточного содержимого ферментов синтеза гистидина (рис. 10).
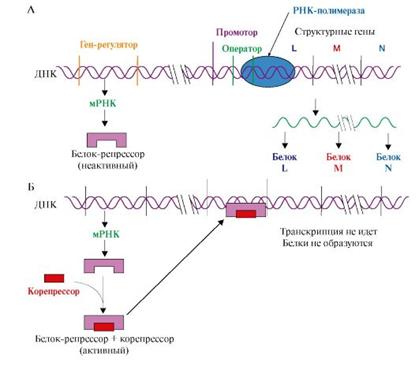
Рис. 10. Оперон, регулируемый по механизму репрессии:
Адаптивная регуляция активности генов у эукариотов обеспечивает изменения скорости транскрипции отдельных генов в ответ на меняющиеся условия внутренней и внешней среды. В клетках многоклеточных организмов часть генов кодирует белки «домашнего хозяйства», которые синтезируются с постоянной скоростью и обеспечивают жизнеспособность клеток. Это - гены ферментов, участвующие в биологическом окислении, синтезе АТФ, образовании компонентов мембран и т.д.
Регуляция у высших организмов отличается от регуляции транскрипции у прокариотов многообразием сигналов, которые контролируют не только начало процесса на молекуле ДНК, но и частоту, с которой он происходит (рис. 11).
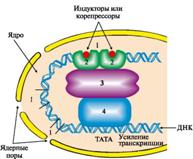
Рис. 11. Адаптивная регуляция транскрипции генов у эукариотов:
1 - регуляторные участки ДНК; 2 - регуляторные белки; 3 - белки-коактиваторы; 4 - РНК-полимеразный комплекс.
Область, обеспечивающая регуляцию экспрессии генов, включает в себя:
Ø промоторный участок;
Ø дополнительные регуляторные последовательности, в которые входят энхансеры, сайленсеры, гормон-чувствительные участки, последовательности GC, СААТ и др.
Белки, связывающиеся с ДНК в этих участках, называют специфическими регуляторными белками. Они влияют на скорость транскрипции генов, взаимодействуя с белками-посредниками или коактиваторами, передающими сигнал на основные транскрипционные факторы и РНК-полимеразу:
ТАТА-участок промотора присоединяет ТАТА-связывающий белок, факторы транскрипции обеспечивают взаимодействие с РНК-полимеразой и определяют стартовую точку транскрипции. Минимальный синтез мРНК становится возможным после связывания РНК-полимеразы с группой дополнительных транскрипционных факторов. Если кроме указанных компонентов с РНК-полимеразным комплексом связываются белки, присоединенные к регуляторным участкам ДНК, то скорость транскрипции меняется. Она возрастает, если это будут белки, взаимодействующие с участками ДНК-энхансерами (усилителями), и снижается, если к транскрипционному комплексу присоединится белок, связывающийся с участком сайленсера (тушитель транскрипции).
Регуляторные зоны ДНК - энхансеры и сайленсеры, гормон-чувствительные участки, специфические регуляторные последовательности (например, GC, CAAT и др.) различны по числу и расположению на молекуле ДНК для разных генов в разных тканях, т.е. являются тканеспецифическими характеристиками. Они могут располагаться за тысячи нуклеотидных пар от стартовой точки транскрипции впереди, после или внутри гена, связывать комплексы белков с метаболитами или гормонами и влиять на конформацию гена.
Индукторами или корепрессорами, стимулирующими присоединение регуляторных белков к ДНК, могут быть:
Ø гормоны,
Ø ионы металлов,
Ø субстраты или продукты метаболических путей.
Определенное значение в регуляции состава и содержания белков имеют посттранскрипционные превращения пре-мРНК в процессе альтернативного сплайсинга, изменение стабильности РНК в разные периоды жизни клетки.
Рис. 6. Инициация белкового синтеза.
Этап элонгации включает три последовательные стадии:
1. связывание аа-тРНКаа в А-центре (требует затраты энергии ГТФ и участия фактора элонгации EF1 - на схеме аа-тРНКаа - Вал-тРНКВал);
2. образование пептидной связи катализирует пептидилтрансфераза, активный центр которой формируется рРНК, входящей в состав большой субъединицы рибосомы;
3. перемещение рибосомы по мРНК на один кодон в направлении от 5'- к 3'-концу идет с использованием энергии ГТФ (транслокация) и при участии фактора EF2, рис. 7.
Рис. 7. Элонгация полипептидной цепи
Терминация трансляции происходит после включения в А-центр одного из стоп кодонов: UAG, UGA, UAA, рис. 8.
Рис. 8. Терминация синтеза белка
Одновременно несколько рибосом могут участвовать в трансляции одной мРНК. Каждая рибосома занимает участок, равный примерно 80 нуклеотидам мРНК. Таким образом, рибосомы располагаются на мРНК с интервалами около 100 нуклеотидов, образуя комплекс, называемый полисомой.
Регуляция активности (экспрессии) генов. Модель работы оперона.
Адаптация организмов к различным воздействиям окружающей среды часто осуществляется путем изменения экспрессии (активности) генов, т.е. изменения скорости транскрипции определенных участков на молекулах ДНК. Этот процесс детально изучен на бактериях.
Адаптивная регуляция активности генов у прокариотов получила объяснение в теории оперона. Согласно этой теории на молекуле ДНК прокариотов присутствуют определенные участки - опероны.
В состав оперона входят:
1. структурные гены - содержат информацию о группе функционально взаимосвязанных белков, которые участвуют в одном и том же метаболическом пути;
2. промотор – расположен в начале оперона, короткая последовательность в несколько десятков нуклеотидов ДНК, с которой специфически связывается фермент РНК-полимераза
3. оператор - участок, с которым происходит связывание белка-репрессора.
4. терминатор – последовательность нуклеотидов, прекращающая транскрипцию.
Участки промотора и оператора частично перекрываются.
Белок-репрессор синтезируется в клетке с постоянной скоростью, его строение кодирует мРНК, транскрибируемая с гена-регулятора, расположенного на некотором расстоянии от оперона, работу которого контролирует его белковый продукт.
Если оперон регулируется по механизму индукции (например, лактозный оперон), то в отсутствии индуктора (лактозы) белок-репрессор связан с оператором. Комплекс белок-репрессор-оператор препятствует связыванию РНК-полимеразы с промотором, и транскрипция структурных генов оперона не идет (рис. 9 А).
Когда концентрация индуктора в клетке возрастет, то он присоединится к белку-репрессору, который имеет центр для связывания индуктора, изменяет его конформацию и снижает сродство к оператору. Комплекс индуктор-белок-репрессор теряет сродство к ДНК и уходит в цитозоль клетки. РНК-полимераза связывается с промотором и транскрибирует структурные гены. Идет синтез белков, закодированных в данном опероне (рис. 9 Б).
Рис. 9. Оперон, регулируемый по механизму индукции (лактозный оперон)
При регуляции оперона по механизму репрессии (например, гистидиновый или триптофановый опероны) белок-репрессор, постоянно синтезируемый в клетках, не имеет сродства к оператору (рис. 10 А). Когда к белку-репрессору присоединится небольшая молекула - корепрессор (гистидин или триптофан, как правило, конечный продукт метаболического пути, ферменты которого закодированы в структурных генах оперона), то в результате конформационных изменений комплекс белка-репрессора с корепрессором приобретает сродство к оператору и прекращает транскрипцию (рис. 10 Б). Так, гистидиновый оперон содержит 10 структурных генов, кодирующих строение ферментов синтеза гистидина. Добавки гистидина в среду выращивания клеток E. Coli вызывают снижение количества, а затем и полное исчезновение из внутриклеточного содержимого ферментов синтеза гистидина (рис. 10).
Рис. 10. Оперон, регулируемый по механизму репрессии:
Адаптивная регуляция активности генов у эукариотов обеспечивает изменения скорости транскрипции отдельных генов в ответ на меняющиеся условия внутренней и внешней среды. В клетках многоклеточных организмов часть генов кодирует белки «домашнего хозяйства», которые синтезируются с постоянной скоростью и обеспечивают жизнеспособность клеток. Это - гены ферментов, участвующие в биологическом окислении, синтезе АТФ, образовании компонентов мембран и т.д.
Регуляция у высших организмов отличается от регуляции транскрипции у прокариотов многообразием сигналов, которые контролируют не только начало процесса на молекуле ДНК, но и частоту, с которой он происходит (рис. 11).
Рис. 11. Адаптивная регуляция транскрипции генов у эукариотов:
1 - регуляторные участки ДНК; 2 - регуляторные белки; 3 - белки-коактиваторы; 4 - РНК-полимеразный комплекс.
Область, обеспечивающая регуляцию экспрессии генов, включает в себя:
Ø промоторный участок;
Ø дополнительные регуляторные последовательности, в которые входят энхансеры, сайленсеры, гормон-чувствительные участки, последовательности GC, СААТ и др.
Белки, связывающиеся с ДНК в этих участках, называют специфическими регуляторными белками. Они влияют на скорость транскрипции генов, взаимодействуя с белками-посредниками или коактиваторами, передающими сигнал на основные транскрипционные факторы и РНК-полимеразу:
ТАТА-участок промотора присоединяет ТАТА-связывающий белок, факторы транскрипции обеспечивают взаимодействие с РНК-полимеразой и определяют стартовую точку транскрипции. Минимальный синтез мРНК становится возможным после связывания РНК-полимеразы с группой дополнительных транскрипционных факторов. Если кроме указанных компонентов с РНК-полимеразным комплексом связываются белки, присоединенные к регуляторным участкам ДНК, то скорость транскрипции меняется. Она возрастает, если это будут белки, взаимодействующие с участками ДНК-энхансерами (усилителями), и снижается, если к транскрипционному комплексу присоединится белок, связывающийся с участком сайленсера (тушитель транскрипции).
Регуляторные зоны ДНК - энхансеры и сайленсеры, гормон-чувствительные участки, специфические регуляторные последовательности (например, GC, CAAT и др.) различны по числу и расположению на молекуле ДНК для разных генов в разных тканях, т.е. являются тканеспецифическими характеристиками. Они могут располагаться за тысячи нуклеотидных пар от стартовой точки транскрипции впереди, после или внутри гена, связывать комплексы белков с метаболитами или гормонами и влиять на конформацию гена.
Индукторами или корепрессорами, стимулирующими присоединение регуляторных белков к ДНК, могут быть:
Ø гормоны,
Ø ионы металлов,
Ø субстраты или продукты метаболических путей.
Определенное значение в регуляции состава и содержания белков имеют посттранскрипционные превращения пре-мРНК в процессе альтернативного сплайсинга, изменение стабильности РНК в разные периоды жизни клетки.