ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 22.11.2023
Просмотров: 154
Скачиваний: 1
Нейросекрет оказывает физиологическое действие через гуморальную среду организма. В связи с этим значительный интерес представляет вопрос о путях его выведения из нейронов ядер гипоталамуса. Микроскопическими исследованиями установлено, что из невронов супраоптических и паравентрикулярных ядер гранулы гомориположительного нейросекрета по длинным аксонам гипоталамо-гипофизарного тракта токами аксоплазмы смещаются в заднюю долю гипофиза. По мере передвижения от места образования до окончания нервных отростков изменяются химические и тинкториальные свойства нейросекрета. По данным отдельных исследователей, нейросекрет может синтезироваться и в терминалях аксонов, лежащих в задней доле гипофиза (Ди-пен). Задняя доля гипофиза является, таким образом, резервуаром для гипоталамического гомориположительного нейросекрета. Отсюда нейросекрет поступает в кровеносное русло. Часть нейро-секреторного вещества может попадать в кровеносное русло и через ликвор третьего желудочка. Кроме того, нейросекреторные клетки посылают свои аксоны к обонятельным ядрам и эпендиме боковых желудочков переднего мозга. Депонированный в задней доле гипофиза, гомориположительный нейросекрет является носителем физиологически высокоактивных гормонов вазопрессина и окситоцина. По мнению некоторых авторов,_окситоцин образуется в паравентрикулярном, а вазопресин в супраоптическом ядрах. Другие же исследователи считают, что, в зависимости от физиологического состояния организма, клетки одного и того же ядра могут синтезировать оба гормона.Р ис. 75. Портальная системасосудов гипофиза у птиц (поГлису):1 — передняя доля гипофиза; 2 — задняя доля гипофиза; 3 — срединное возвышение.Гипоталамический нейросекрет поступает также в переднюю долю гипофиза и контролирует ее гормональную функцию. Однако сюда он поступает из подбугорья не по нервным окончаниям, а с током крови, циркулирующей по сосудам портальной системы. Гуморальный путь тра-нспортировки вырабатываемых клетками гипоталамуса веществ в переднюю долю гипофиза доказывается прямыми экспериментами. У уток, например, портальные вены из гипоталамуса в аденогипофиз проходят обособленно от гипофизарной ножки (рис. 4). Бенуа и Ассенмахер указывают, что перерезка только одной ножки не оказывает существенного влияния на гормональную функцию гипофиза. Перерезка же портальной системы сосудов, при

сохранении целостности гипофизной ножки, приводит к угнетению физиологической активности передней доли гипофиза. Показано также, что кровь, взятая из портальных сосудов, стимулирует гипофиз, тогда как кровь из сонной артерии этими свойствами не обладает. Эти эксперименты показывают, что из гипоталамуса в переднюю долю гипофиза действительно поступают вещества, которые активируют его гормональную деятельность. Однако природа этих веществ изучена еще недостаточно.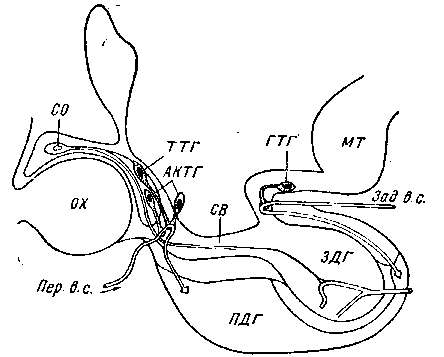
Рис. 5. Схема локализации зон в гипоталамусе собаки, ответственных
за рефляцию тиреотропной (ТТГ), адренокортикотропной (АКТГ) и гонадо-
троной (ГТГ) функции гипофиза (по Хьюму):
Пер. в. с. — передние воротные сосуды; Зад. в. с.—задние воротные сосуды; МТ —
маммилярпос тело; X — хиазма; СО — супраоптическое ядро; ПДГ — передняя доля
гипофиза; ЗДГ — задняя доля гипофиза
Микроскопическими исследованиями установлено, что во внутреннем слое срединного возвышения амиэлиновые волокна ту-бероинфундибулярного пучка, берущие начало от клеток ядер серого бугра, своими окончаниями вступают в связь с расположенными здесь короткими петлями и клубочками первичных капилляров портальной системы (см. рис. 2).
В этих вазоневральных синапсах отростки нервных клеток гипоталамуса отдают в кровь портальной системы секреторное вещество, которое поступает затем в паренхиму передней доли гипофиза. Этот нейросекрет, выделяемый ядрами серого бугра, однако, не окрашивается по Гомори. Отдельные исследователи отождествляли его с медиаторами нервных клеток (ацетилхоли-ном и норадреналином). В настоящее время эта теория уже почти совсем не имеет сторонников. Исследования показали, что нейро-секрсторное вещество от обычных метаболитов нервных клеток отличается более высокой энзиматической устойчивостью и способностью оказывать физиологическое действие на значительном расстоянии от места своего образования в гипоталамусе. Значительный интерес представляет тот факт, что при электролитическом повреждении отдельных участков срединного возвышения в передней доле гипофиза подавляется образование и выведение
в кровь не всех, а только какого-либо одного гормона, активирующего функцию строго определенной периферической эндокринной железы (рис. 5). На этом основании считают, что отдельные нервные волокна от различных клеток гипоталамуса несут в срединное возвышение и отдают в кровь портальной системы не одно, а несколько особых веществ или нейрогуморов (Каррато и др.), которые реализуют различные функции передней доли гипофиза (гонадотропную, тиреотропную и адренокор-тикогропную). Возможно, различные нейрогуморы сосудами портальной системы проецируются на определенные зоны паренхимы передней доли гипофиза.Как уже отмечалось, значение гомориположительного нейро-секрета в регуляции гормональной функции гипофиза изучено еще недостаточно. Обычными методами окрашивания его не удается выявить в туберо-гипофизарном пути и в крови портальной системы. Вместе с тем вокруг капилляров, в их эндотелии и между секреторными клетками передней доли гипофиза часто накапливается значительное количество гранул, которые проявляют такие же реакции, как и гипоталамический неиросекрет. При избыточном введении в организм поваренной соли происходит быстрое выведение нейросекрета из задней доли гипофиза, супраоптического и паравентрикулярного ядер. В этих условиях нейросекретом обогащаются и секреторные клетки передней доли гипофиза (Войткевич и др.). Отдельные исследователи допускают, что в осуществлении связи гипоталамуса с гипофизом важную роль могут играть вазопресин и окситоцин, носителями которых является неиросекрет паравентрикулярного и супраоптического ядер (Мартини и др.). Следует, однако, отметить, что методом хроматографии Саффрон удалось отделить в экстрактах задней доли гипофиза от вазопресина и окситоцина вещество, которое активирует адренокортикотропную функцию передней доли гипофиза.Приведенные данные показывают, что вопрос о природе нейросекрета, ответственного за регуляцию функции гипофиза, еще нуждается в дальнейшей разработке. Тем не менее, обширный фактический материал указывает на ведущее значение в гнпота-ламическом контроле функции эндокринной системы васкулярных связей. Срединное возвышение серого бугра гипоталамуса и является тем участком, во внутреннем слое которого через посредство вазоневральных синапсов осуществляется передача влияний с гипоталамуса на переднюю долю гипофиза.
Значение гипоталамуса в регуляции функции эндокринной системы. Гипоталамус принимает участие в нервной и гуморальной регуляции физиологических функций организма. Особенно велико его значение в контроле гормональной деятельности эндокринной системы. Прежде всего гипоталамус сам продуцирует вещества, которые гуморальным путем влияют на отдельные функции организма. Уже отмечалось, что нейроны супраоптического и паравентрикулярного ядер подбугорья синтезируют неиросекрет, который перемещается по нервным отросткам гипоталамо-гипофизарного тракта и аккумулируется в задней доле гипофиза. Этот нейросекрет является носителем физиологически высокоактивных гормонов вазопресина и окситоцина.Клинические наблюдения и многочисленные экспериментальные исследования последних лет показывают, что гипоталамус оказывает доминирующее влияние на гормональную деятельность передней доли гипофиза и через нее на многие периферические железы внутренней секреции. Этот вывод основан прежде всего на экспериментах по нарушению анатомической связи между гипоталамусом и гипофизом. Так, при перерезке у кроликов (Гаррис), кур (Ширм и Налбандон), уток (Бенуа и Ассен-махер) и других животных гипофизарной ножки резко понижается выделение гипофизом в кровь кринотропных гормонов, активирующих функцию половых желез, коры надпочечников и щитовидной железы. При нарушении связи между гипофизом и гипоталамусом периферические железы переходят в состояние физиологической депрессии. Эта операция особенно сильно отражается на функциональном состоянии половых желез. Если после перерезки ножки портальные сосуды регенерируют и восстанавливается транспортировка из гипоталамуса нейросекрста, то тогда опять нормализуется работа передней доли гипофиза и периферических желез.Представления о механизмах передачи регулирующих влияний с гипоталамуса на гипофиз за короткую историю разработки этой важной проблемы современной эндокринологии претерпели существенные изменения. На первых этапах ее разработки многие исследователи считали, что влияние гипоталамуса на гипофиз осуществляется через посредство парасимпатической и симпатической нервной системы. Поскольку, однако, прямая нервная связь между ними, по-видимому, отсутствует, то Шаррер уже давно высказал предположение, что гипоталамическая регуляция гормональных функций гипофиза осуществляется преимущественно гуморальным путем при участии нейросекрета. Это положение в дальнейшем подтвердилось не только в опытах с перерезкой портальных сосудов, но и в экспериментах по трансплантации гипофиза в различные органы. При пересадке его гипофизэктомированным животным в почку или в височную долю (Гаррис и Якобсон) гипофиз приживляется, васкуляризируется, но в этих условиях угнетается его гормональная деятельность. Если же этот гипофиз затем подсадить в область срединного возвышения, то после врастания портальных сосудов его гормональная деятельность быстро восстанавливается. Такой же результат получен при совместной инкубации вне организма гипофиза с фрагментами гипоталамуса или при добавлении в культуру экстракта из срединного возвышения подбугорья.