ВУЗ: Казахский национальный медицинский университет им. С.Д. Асфендиярова
Категория: Шпаргалка
Дисциплина: Биология
Добавлен: 05.02.2019
Просмотров: 2073
Скачиваний: 50
Апоптоз сатыларының мәні, сызбасын сызу және медициналық маңызын түсіндіру.
Апоптоз – жасушаның табиғи, генетикалық бағдарламаланған тіршілігін жою үрдісі.
Апоптоз бағдарламасын іске қосатын жағдайларға байланысты апоптоздың екі типін ажыратуға болады: а)“ішкі” апоптоз ; б)“бұйрық” бойынша апоптоз
а)“Ішкі” апоптозды іске қосатын себептер:
Хромосоманың репарацияланбайтын зақымдалулары; Жасушаішілік органоидтардың және мембраналардың (әсіресе митохондрия) зақымдалуы.
Бұл зақымдалуларды тудыратын факторлар:
Ішкі ( күшті тотықтырғыштар,азот оксиді, супероксидті радикалдар)
Сыртқы (иондаушы сәулелер, t өзгеруі, химиялық қосылыстар)
Апоптоздың бұл жағдайда негізгі қызметі – құрылысы не қызметі зақымдалған жасушаны жою.
б) “Бұйрық” бойынша апоптозды іске қосатын факторлар:
1. Негативті сигнал, мысалы контактылы тежелуде, кадгериндерден келетін сигнал. Бұл жағдайда бөлінуші жасушалар өте тығыз орналасуы, тек олардың бөлінуін тоқтатып ғана қоймай, апоптозға ұшырауы мүмкін.
2. Оң сигналдардың әсерінің тоқтауы :
-митогендер әсерінің болмауы (цитокин не өсу факторлары)
-бекіну беткейінен ажырау (интегрин сигналы)
Бұл жасушада апоптоздық белок р53 мөлшерінің шектік деңгейден артуына алып келеді.
Апоптоз үрдісіндегі негізгі ферменттер – цитоплазмалық протеазалар – каспазалар болып табылады.Каспазалар барлық жасушалардың цитоплазмасында активсіз күйінде – прокаспазалар түрінде болады.
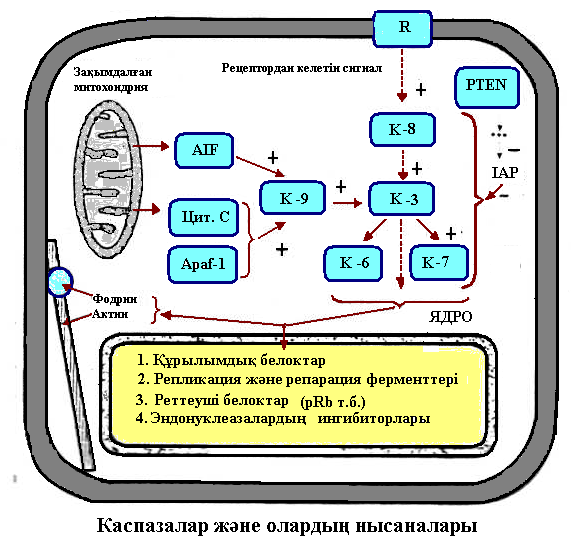
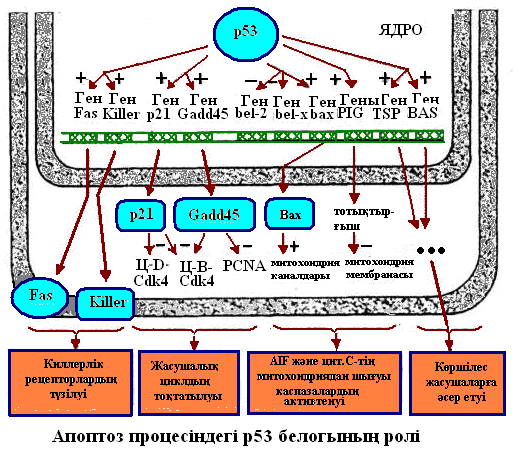
Митохондрия каналдарын ашатын және жабатын белоктарды кодтайтын гендер р53 белогымен реттеледі.
Апоптоз үрдісі тірі ағза тіршілігінде өте маңызды роль атқарады:
онтогенез барысында (саусақ аралық жарғақтың жойылуы, ішек, тамыр қуыстарының қалыптасуы); жасушалардың жаңаруы (қан жасушалары, тері, ішкі мүше эпителийлері);
Иммундық жүйенің қызметінде (антиген әсерінен кейін лимфоциттердің ұзақ уақыт антиген болмаған жағдайда жойылуы және т.б.);
Сонымен бірге апоптоз үрдісінің бұзылуы: ісіктік жасушалардың өсуін шектеудің бұзылуына, ісіктік өсуге; аутоиммунды аурулар;дегенеративті ауруларға алып келеді.
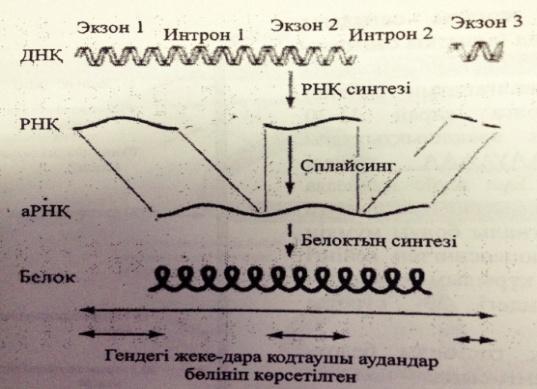
А-РНҚ процессингінің механизімін сызу түсіндіру
Гетерогендік ядролық РНҚ бірқатар құрылымдық өзгерістерге ұшырап, функциональды активті жетілген а-РНК-ға айналып цитоплазмаға белок синтезделетін жерге жеткізілуі тиіс. Бұл-“ПРоцессинг” деп аталады.
Процессинг механизмі:
1)Қалпақ немесе кэп жалғануы . Кэп жалғану процессі-пре-а-РНК-ның 5-ұшына метилденген гуанозиннің жалғануы
2) Полиаденилдік Құйрықтың жалғануы .
3) РНК –сплайсингі.

Гендік мутациялар ДНҚ молекуласының құрылысындағы өзгерістермен сипатталады. Олар 2-ге бөлінеді:
1)репликация қателіктері-нуклеотид жұптарының алмасуы;
а)егер пуриндік негіздер пуриндік негіздерге, пиримидиндік негіздер пиримидиндік негіздерге ауысса транзиция.
б) егер пурин пиримидинге немесе керісіеше ауысса трансверция.
2) оқылу ретінің жылжуы- нуклеотидтердің түсіп қалуы немесе қосылуы;
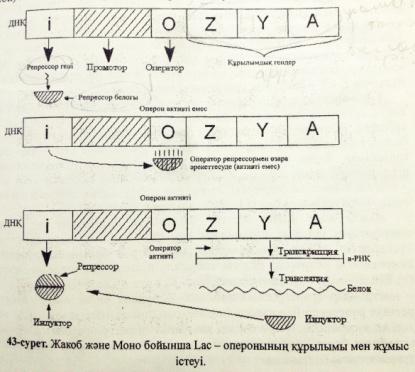
Ген белсенділігінің оперондық моделінің сызбасын сызу және түсіндіру
Алғаш рет Ф. Жакоб және Ж. Моно прокариоттарда гендер активтілігінің реттелуін зерттеу мақсатында ішек таяқшасында β-галактозидаза ферментінің синтезі реттелуіне тәжірибе жүргізді. Тәжірибе нәтижесінде фермент синтезделудің индукциясв мен репрессиясының механизмдерін түсіндіретін “оперон моделін” ұсынды.
Жакоб және Мононың бұл моделі бойынша оперон құрамында:
1. Бірнеше белок-ферменттердің синтезін анықтайтын қатарласа орналасқан құрылымдық гендер болады .
2.Осы гендердің активтілігін бақылайтын реттеуші немесе оператор гені болады.
Оперондағы құрылымдық гендердің ерекшелігі –олар оперонда қатарласа орналасқан кластерлік гендерді құрайды . Осы кластерлік гендерден тұтас бір ғана полицистронды РНҚ түзіледі.Лактозалық оперондағы құрылымдық гендер 3-цистроннан тұрады:Z,Y,A.Бұл гендердің өнімі –ферменттер лактоза қантын ыдыратуға қатысады.
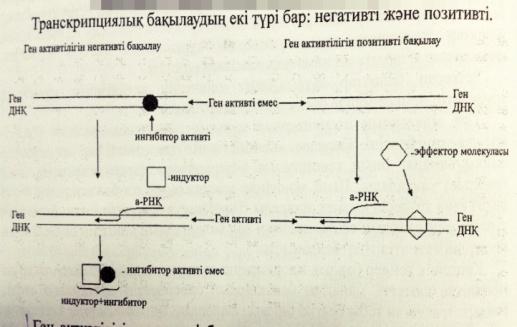
Ген белсенділігінің позитивті реттелуінің сызбасын сызу және түсіндіру
Позитивтік бақылану механизмі нақты фактор бар кезде -активтілікті күшейтіп РНҚ-полимеразаның промоторға жалғануы мен а-РНҚ-ның синтезделуін жылдамдатады. Позитивті бақылаудың мәні мынада-жасушада глюкоза мен галактоза бар кезде LAC-оперон активті емес сондықтан LAC-а-РНҚ –ның синтезі жүрмейді β-галактозидаза ферменті де синтезделмейді. Лактозалық i-репрессорын активсіздендіру үшін ортада тек глюкозаның болғаны жеткіліксіз.Глюкозаға қосымша элемент циклдық АМФ-катаболизмдік активтендіруші белок КАБ болуы қажет .
Циклдық
АМФ-тың кішігірім молекулалары ішек
таяқшасынан басқа көптеген бактерияларда
да кездеседі, олардың синтезделуі
аденил-циклаза ферментінің көмегімен
жүреді.
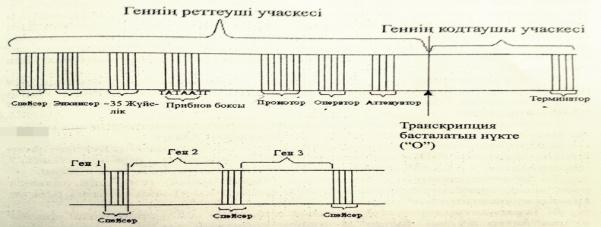
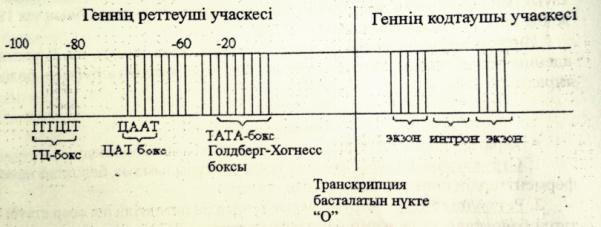
Геннің молекулалық деңгейдегі құрылысын және қызметтерін сызу және түсіндіру.
Ген-күрделі құрылымдық функциональдық бірлік болып табылады.Ол екі бөліктен тұрады:
1)реттеуші учаске,2)ақпараты бар кодтаушы учаске.
Ген-полипептидтік тізбек. Бір ген-бірфермент,бір цистрон-бірполипептидтік тізбек.
Прокариоттар генінің молекулалық құрылымы
Эукариоттар генінің молекулалық құрылымы
Промотор-транскрипция кезінде РНҚ полимеразамен байланысады.
Оператор-реттеуші белоктардың байланысатын арнайы нуклеотидтер жүйесі.
Транскрипция-басталатын нүкте немесе «О»сайты.
Энхансер-транскрипция жылдамдығын 200 есеге дейін арттырады.
Сайлансер-транскрипция жылдамдығын төмендетеді.
Прибнов-бокс-промотор учаскесі ішіндегі арнайы нуклеотидтер жүйесі.
Аттенуатор-транскрипцияның аяқталуы жайлы белгі беретін арнайы жүйесі.
Терминатор-транскрипцияның аяқталғандығы жайлы сигнал береді.
Генетикалық кодтың қасиеттерінің сызбасын сызу және түсіндіру
Тұқым қуалау ақпараты ДНҚ молекуласында 4түрлі нуклеотидтің нақты бір жүйемен кезектесіп орналасуы түрінде жазылған(кодталған). Кодтаудың негізгі принциптері Ф.Крик және оның қызметкерлерімен құрастырылып, «генетикалық код»деп аталған. Генетикалық код 1965 жылы толық анықталды.
Генетикалық кодтың қасиеттері:
-триплеттілігі -қатарынан орналасқан 3нуклеотид бір кодон (триплет)құрап, бір аминқышқылын анықтайды
-үздіксіздігі -кодондар үздіксіз орналасқан
- қайта жабылмайтындығы -кодондар бірі бірін жаппайды, әр нуклеотид тек бір кодонның құрамына кіреді
-коллинеарлығы -кодондардың реті белоктағы аминқышқылдардың ретіне сай
-артықтылығы - бір аминқышқылды 1-6 жуық кодон анықтауы мүмкін, өйткені жалпы кодондар саны 64, ал аминқышқылдар саны – 20
-жүйелілігі - кодондағы нуклеотидтердің ақпараттық маңыздылығының бірдей болмауы: 1-2 орындағы нуклеотидтер маңызды болып саналады, ал 3 нуклеотид әртүрлі болуы мүмкін және автоматты түрде оқылады
-арнайылылығы - әр кодон тек белгілі бір аминқышқылын анықтайды
-универсальдығы - барлық тірі ағзаларға (бірқатар ерекшеліктерінен басқа) генетикалық код бірдей
Геннің трансляция үдерісін сызу және түсіндіру
Трансляция – полипептидтік тізбектің матрицалық синтезделу процессі. Бұл процесс цитоплазмада рибосомалар, тРНҚ және аРНҚ қатысуымен жүзеге асырылады. Кез келген матрицалық синтез сияқты, ол иницияция, элонгация және терминация сатыларынан тұрады.
Геномдық мутациялар мейоз немесе митоздың бұзылуының нетижесінде хромосомалардың диплоидтық жиынтығының санының өзгеруінен болатын мутациялар. Оларға полиплоидия, гаплоидия, гетероплоидия (анеуплоидия) жатады.
Полиплоидия хромосоманың диплоидтық санының гаплоидтық жиынтығына еселеніп артуын айтамыз. (2n+n;2n+2n;2n+3n;)
Гаплоидия хромосомалардың тек гаплоидты жиынтығының болуы.(n)
Гетероплоидия (анеуплоидия) диплоидтық жиынтықта жеке хромосомалар санының өзгеру.
Нулисомия-(2n-2), моносомия-(2n-1), трисомия-(2n+1), тетрасомия-(2n+2).
ДНҚ репликациясының принциптерінің сызбасын сызу және түсіндіру.
ДНҚ молекуласының екі еселену процесі репликация деп аталады. Репликацияның бірлігі-репликон,өздігінен репликацияланатын генетикалық элемент.Оның құрамында ДНҚ репликациясының инициациялық учаскесі және репликация процесіне қатынасатын белоктардың синтезін бақылайтын гендер орналасқан.Репликация аса күрделі биохимиялық процесс.Бұл процеске генетикалық ақпараттың өте дәлдікпен көшірілуін,ДНҚ тізбектеріндегі қателіктерді түзетіп,алмастыруды және рекомбинациясын қамтамасыз ететін көптеген белоктар қатынасады.
Репликация процесі мына принциптің негіізінде іске асады:
1)жартылай консервативтік
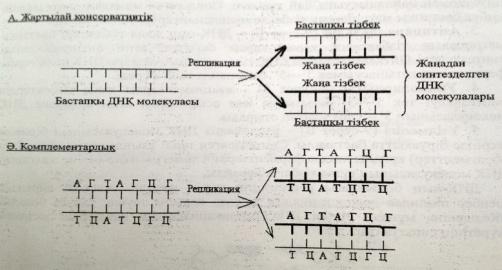
2)комплементарлы
3)Антипараллельдік және униполярлық
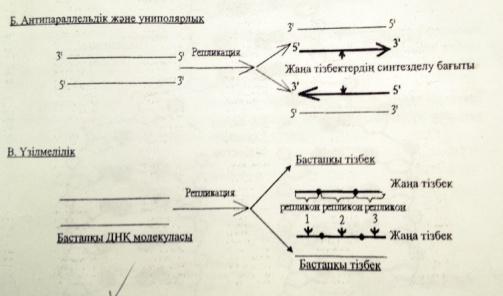
4)Үзілмелілік
1) Жартылай консервативті-ДНҚ-ның бастапқы тізбектерінің әрқайсысы жаңа тізбек түзілу үшін матрица болып табылады.
Репликациядан кейінгі әр ДНҚ молекуласындағы екі тізбектің біреуі бастапқы матрицалық тізбек болса,екіншісі –жаңадан синтезделетін тізбек.Олай болса,алғашқы аналық жасушаның бөліну нәтижесінде түзілген әр жас жасушадағы ДНҚ молекуласы бір бастапқы матрицалық тізбектен және бір жаңадан синтезделетін тізбектен тұрады.
2)Комплементарлық-ДНҚ молекуласының жаңа тізбегі комплементарлық принципке негізделіп адениннің тиминмен,гуаниннің цитозинмен байланысуына сай түзіледі.Сондықтан жаңадан синтезделген тізбек бастапқы матрицалық тізбекке комплементарлы болып келеді.
3)Антипараллельдік-ДНҚ-ның жаңа тізбектері бастапқы матрицалық тізбектерге қарама-қарсы бағытта,яғни антипараллельді синтезделеді.Өйткені бастапқы ДНҚ-дагы ақпараттың оқылуы ДНҚ-полимераза ферментінің қатынасуымен 3 1 5 1 бағытында ғана жүреді.
4)Униполярлығы-комплементарлы жаңа тізбектердің синтезделуі тек 5 1 31 бағытында іске асырылады,яғни әрдайым ДНҚ молекуласының 31 ұшы ұзарып отырады.
5)Үзілмелігі-репликация ДНҚ молекуласының бірнеше жерінде бір уакытта басталады.Синтезделген түрлі ұзындықтағы кесіндіілер арнайы лигаза ферменттерінің көмегімен бір-біріне жалғанып ДНҚ молекуласының тұтас тізбегін құрайды.
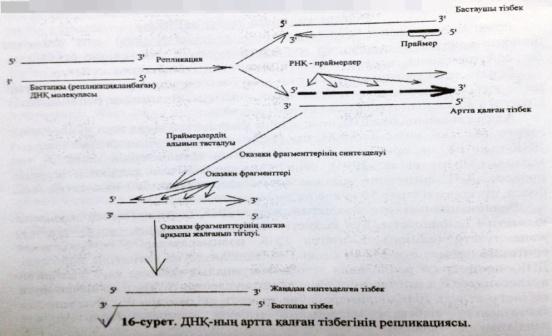
ДНҚ-ның лидерлік және ілесуші тізбектерінің репликациясының сызбасын сызу және түсіндіру.
ДНҚ синтезі спиральдың 51-ұшына қарай жүреді.Репликативтік ашаның бір спиралінде ДНҚ үзіліссіз тізбек синтезделінеді оны лидерлік тізбек,ал екіншісінде үзіліп-үзіліп жүреді,оны ілесуші тізбек деп атайды.
ДНҚ-ның ширатылған тізбектері тарқатылып репликация басталатын учаскеде «репликациялық айыр»
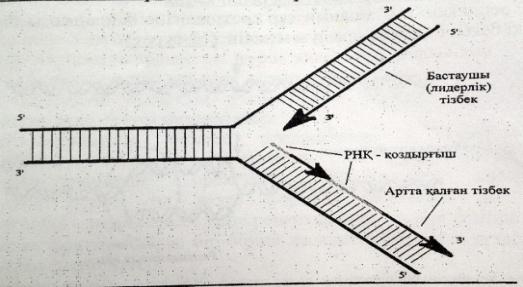
деп аталатын арнайы құрылым түзіледі.
Репликация кезінде ДНҚ синтезделу бвғытының қос тізбектің таралу бағытымен сәйкес келуі тек жаңа синтезделген бастаушы тізбекте көрінеді.Екінші,артта қалған тізбек үзік-үзік болыпқысқа Оказаки ферметтері трінде синтезделеді.Нәт. тізбектің екеуі де 51 → 31бағытында ұзарады.
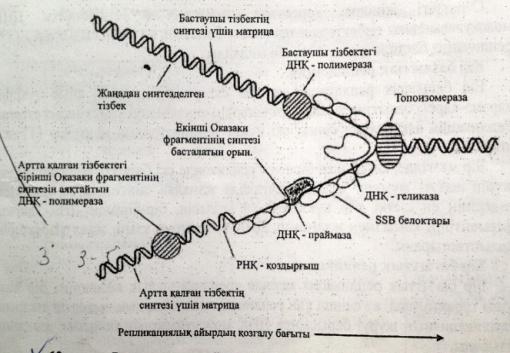
Репликазаның негізгі ферменттері:1.ДНҚ-полимераза-тізбекті синтездейді.2.Топоизомераза-«репликативтік айырдың» алдындағы ДНҚ-ның аса жоғары ширатылған жерлерін босатады.3.Хеликаза-ДНҚ тізбектерін ажыратады.4.SSB-белоктар-ДНҚ-ның ажыраған тізбектерін тұрақтандырады.5.Лигаза-ДНҚ фрагменттерін жалғап қосады.6.РНҚ-праймаза-ДНҚ-полимеразаға керекті РНҚ-бастауыштарды синтездейді.7.ДНҚ-геликаза-комплементарлы нуклеотидтердің арасындағы сутекті байланыстарды үзеді.
ДНҚ молекуласының жарықтылық, қараңғылық және репликациядан кейінгі репарацияның сызбасын сызу және маңызын түсіндіру.
ДНҚ репарациясының типтері:
1. Жарықтық репарация немесе фотореактивация.
1.Қалыпты ДНҚ молекуласы-----Ультракүлгін жарығымен сәулелендіру-----2.Мутантты ДНҚ молекуласы-пиримидиндік димерлердің түзілуі-----Көзге көрінетін жарықтың әсері-----3.Фотолиаза ферментінің синтезі-----4.Димерлердің ажыратылуы-----5.ДНҚ құрылысының бастапқы қалпына қайта келуі.
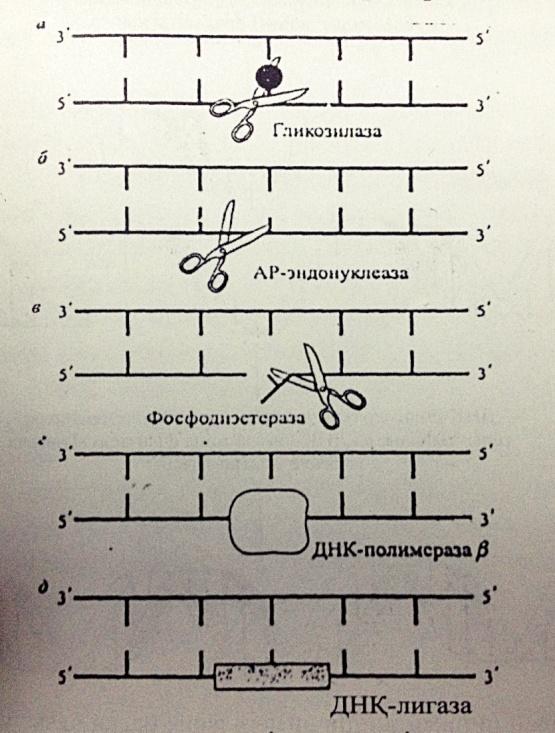
2. Эксцизиялық немесе қараңғылық репарация- жасушадағы жарықтың қатысуынсыз ақ жоя алады. ДНҚ-ның бұзылған учаскесі арнайы ферменттер тобының қатынасуымен кесіп алынып тасталады, оның орнына қалыты нуклеотидтер жалғанады. Келесі сатылардан тұрады:
а)эндонуклеаза ферментінің көмегімен зақымдалған ДНҚ-ны анықтау; б)эндонуклеаза көмегімен зақымдалған аймақты кесіп алу; в)полимераза көмегімен жаңа тізбекті қалыпқа сай синтездеу; г)полинуклеотидлигаза ферментінің әсерінен жаңа түзілген ДНҚ аймағының қосылуы. Мұны эксцизиялық, яғни ”кесумен” жүретін деп атайды.
3. Репликациядан кейінгі репарация- егер фотореактивация және эксцизиялық репарация белгілі бір себептерге байланысты жүрмесе, онда тізбектегі зақымдалу дұрысталмайды және жаңа синтезделген тізбекте бос қуыс қалады. Бұл қуыс екінші зақымдалған ДНҚ тізбегінен синтезделген жаңа комплементарлы аймақпен толтырылады. Бір зақымдалмаған тізбек барлық уақытта қалып ретінде қажет.
Жасушаның генетикалық материалының ұйымдасуының әртүрлі деңгейлерінің механизмдерінің сызбасын сызу және түсіндіру
Жасушаның генетикалық материалының ұйымдасуының деңгейлері:
-
ДНҚ тығыздалуының бірінші деңгейі – нуклеосомалық жіпше(Фибрилла) – жуандығы 10нм, сыртын орап орналасатын ДНҚ-ның ұзындығы 146ж.н. тығыздалу коэффиценті – 6-7
-
Екінші деңгей – жуандығы 30-нм-лік соленоид жіпшесі; тығыздалу коэфф.- 40
-
Үшінші деңгей – ілмектік домен (хромомера); 60мың ж.н. қамтитын ұзындығы 0,2-0,3мкм, тығ.коэфф.-680
-
Хромосомалық төртінші деңгей; тығ-н хромонемалар ұзындығы 0,1-0,2мкм жуан жіпшелері түзейді, олар жарық микроскопы астында көрінеді, тығыздалу коэфф. - 12×104
-
Хроматидалық және хромосомалық деңгей жарық микроскопы астында анық корінетін хроматин құрылымының жоғары деңгейі болып табылады.

Жасушаның генетикалық материалының митоздық циклда өзгеруінің сызбасын сызу және түсіндіру
Митоз – сомалық жасушаның негізгі бөліну әдісі. Митоз бірінен кейін бірі жүретін 4фазадан тұрады: профаза,метафаза, анафаза,телофаза. Митоз – бұл хроматидтер бір-бірінен ажырап, екі жас жасуша арасында бірдей бөлінетін ядроның бөліну процесі.
-
Профаза – хромасомалар ширатылып,жуандап,қысқарады.Әр хромасома 2хроматидтен тұрады. Олар центромералары арқ.байланысады. Ядрошықтар жойылып,ядро қабығы еріп кетеді. Хромасомалар цитоплазмада бос күйінде қалады.Осы кезде центроильдер жасушаның полюстеріне ажырап,ахроматин жіпшелері пайда болып,бөліну ұршығы қалыптасады.
-
Метафаза – хромосомалар экватор жазықтығында орналасады.Әр хромосома кинетохор арқылы бөліну ұршығының жіптеріне бекінген ұзын бойынан екі хроматидтерге ажырайды.
-
Анафаза – хромосомалар полюстерге тартылуы,жеткенде екі бірдей толық хромосомалар жиынтығын құрайды.
-
Телофаза – жаңа ядрошық пайда болады.Хр-р деспиральданады,жіңішкеріп ұзарады,ахроматин жіп-і жойылады.Артынша цитокинез жүріп,жасуша екіге бөлінеді.
Жасушаның мейоздық бөліну үрдісі.н және оның генетикалық маңызының сызбасын сызу және түсіндіру
Мейоз-жыныс бездерінде ерекше сомалық жасушалардың бөлінуі.Нәтижесінде түзілген гаметаларда хромосома жиынтығы гаплоидты болады. Мейоз бірінші және екінші мейоздық бөлінулерден тұрады. Әр бөліну 4-кезеңнен тұрады.Бірінші мейоз:
-Профаза1:
1.Лептотена-хромосомалар ширатылады, жуандайды қысқарады ,микроскоппен көрінеді. Генетикалық материал-2n4c.
2.Зиготена-гомологтық жұп хромосомалар жақындасып ұзынынан бір-бірімен беттеседі.
3.Пахитена-гомологтық хромосомалар қосақталған жұптар құрап биваленттер түзеді. Әр бивалент 4хроматиттерден тұрады Генетикалық материал-2n4c. Коньюгацияланған хромосомалар айқасып, сәйкес бөліктермен алмасады ,кроссинговер жүреді.
4.Диплотена-жұптасқан гомологтық хромосомаларда хромотидтер центромера бөлігінде бір-бірімен ажырай бастайды бірақ айқасқан бөліктер хиазмалар арқылы байланыс сақтайды .
5.Диакинез- хромосомалар қатты ширатылады ядро қабығы ериді ядрошық жойылады бөліну ұршығы түзіледі.
-Метафаза1-биваленттер экватор жазықтығына орналасып центромераларымен бөліну ұршығымен байланысады.
-Анафаза1-тұтас хромосомалар 2жақ полюске қарай тартылады. Бұл кездейсоқ процесс Генетикалық материал-n2c.
-Телофаза1-тек хромосомалар ширатылған түрде сақталады.
Екінші мейоз.
Профаза2-өте қысқа болады хромосомалар ширатылған түрде.
Метафаза2-хромосома экватор жазықтығында орналасады.(n2c).
Анафаза2-бір-бірінен ажыраған хроматидтер қарама-қарсы полюстерге тартылады.(nc)
Телофаза2-цитокинез аяқталғанда гаплоидты жиынтығы бар(nc) жыныс жасушалар түзіледі. Маңызы-гаплоидты жиынтығы бар жыныс жасушалары түзіледі.Ұрықтану кезінде екі гаметаның ядросы қосылып зигота түзеді. Егер гаметалардағы хромосомалар саны кемімесе ұрықтану нәтижесінде олардың саны әр ұрпақ сайын екі есе артады. Мейоз кезінде кроссинговер және гомлогтық хромосомалардың кездейсоқ ажырауы нәтижесінде генетикалық материалдың рекомбинациясы жүреді.
Жасушаның генетикалық материалының өзгеру деңгейіне байл. Мутациялық өзгергіштік деп гендер мен хромосомадағы тұрақты өзгерістер нәтижесінде қалыптасқан өзгергіштікті атайды. Генетикалық матириалдың өзгеруіне байланысты мутациялар геномдық, хромосомалық, гендік болып жіктеледі.
-
Геномдық мутациялар мейоз немесе митоздың бұзылуының нетижесінде хромосомалардың диплоидтық жиынтығының санының өзгеруінен болатын мутациялар. Оларға полиплоидия, гаплоидия, гетероплоидия (анеуплоидия) жатады.
Полиплоидия хромосоманың диплоидтық санының гаплоидтық жиынтығына еселеніп артуын айтамыз. (2n+n;2n+2n;2n+3n;)
Гаплоидия хромосомалардың тек гаплоидты жиынтығының болуы.(n)
Гетероплоидия (анеуплоидия) диплоидтық жиынтықта жеке хромосомалар санының өзгеру.
Нулисомия-(2n-2), моносомия-(2n-1), трисомия-(2n+1), тетрасомия-(2n+2).
-
Хромосомалық мутациялар хромосома құрылысының өзгеруімен. Олар хромосома ішілік, хромосома аралық болып бөлінеді.
-
хромосома ішілікке:
Делеция(жетіспеушілік)- хромосома бөлігінің түсіп қалуы.
Дупликация- хромососма бөлігінің екі еселенуі.
Инверсия- хромосома бөлігінің үзіліп 1800 бұрылып сол хромосомадағы орнына қайта жалғануы; перицентрлік-центромера аймағында, парацентрлік-хромосоманың бір иығында жүріп, центромерадан алшақ байқалады;
-
хромосома аралыққа:
Транслокация- гомологтық емес хромосомалардың учаскелерімен алмасуы; Оның бірнеше түрі бар: Реципрокты- гомологтық емес хромосомалар бір-бірімен бөліктерімен алмасады;
Реципрокты емес- бір хромосома екінші гомологты емес хромосоманың бөлігін жалғастырып өз көлемін ұзартады.
Робертсондық транслокация- гомологты емес екі акроцентрлік хромосомалар ұзын иықтарымен центромерлік аймақтары арқылыжалғанады.
-
Гендік мутациялар ДНҚ молекуласының құрылысындағы өзгерістермен сипатталады. Олар 2-ге бөлінеді:
1)репликация қателіктері-нуклеотид жұптарының алмасуы;
а)егер пуриндік негіздер пуриндік негіздерге, пиримидиндік негіздер пиримидиндік негіздерге ауысса транзиция.
б) егер пурин пиримидинге немесе керісіеше ауысса трансверция.
2) оқылу ретінің жылжуы- нуклеотидтердің түсіп қалуы немесе қосылуы;
Қатерлі ісік жасушаларының қасиеттерін атап оларды көрсету.
Қатерлі ісіктер-жер бетіндегі халықтың,әсіресе жасы ұлғайған адамдар арасындағы өлімнің негізгі себертерінің бірі болып табылады.Қалыпты жағдайда жасушалардың бөліну процесі олардың көбеюін жылдамдатып немесе тежеп және өзара бірін-бірі теңестіріп отыратын екі топ факторлармен бақыланады.Рак жасушаларының негізгі қасиеті тоқтаусыз бөлінуге қабілеттілігі,өйткені оларда жасушаның көбеюін тежейтін механизмдер мен факторлар болмайды.Қалыпты жасушаның трансформацияланған жасушаға айналу процесі онкогенез не канцерогенез деп аталады.Рак полиэтиологиялық не көпфакторлы ауруларға жатады,ерекшелігі патологияның себебін анықтау мүмкін болмайды.Жасушалардың ісіктік трансформациясына себеп болатын канцерогендік факторларға иондаушы сәулелену,химиялық қосылыстар,ісікті туғызатын вирустар.
Иондаушы сәулелену-радиацияның жоғары дозасының әсеріне көпшілік жағдайда радиация көзімен тікелей байланысы бар АЭС-тің кәсіпқой мамандары,рентгенолог және радиологтар,радиоактивті сәулеленумен емделетін адамдар,атом бомбасының жарылыман тап болған адамдар ұшырайды.
Химиялық қосылыстар-жеке дара ісіктік жасушалардың өсуі мен бөлінуін жылдамдатуға қабілетті келеді.Оларға төртхлорды көміртегі,метилхолантрен,бензантрацен жатады.Оларды канцерогендік промоторлар деп атайды.Мұндай жекелеген ісіктік жасушалар қалыпты жасушалармен қоршалып тқрғандықтан олардың тоқтату әрекетін жеңе алмай,ұзақ уақыт,жылдар бойы жасырын күйде болып қатерлі ісік жасушаларына трансформацияланбайды.
Ісіктік вирустар-вирустардың генетикалық материалы ДНҚ не РНҚ молекуласы түрінде болады.Кейбір вирустардың геномында онкогендік активтілік көрсететін гендер болады.Ісіктің вирустары жасушаға еніп,оның ДНҚ-на қосылып иелік жасуша геномы және вирус геномынан тұратын рекомбинантты ДНҚ молекуласының құрамында қызмет атқаруға қабілетті келеді.Жасушалардың ісіктік трансформациясында генетикалық факторлардың негізгі роль атқаратынын растайтын мынадай нақты мәліметтер бар:1)адамның қандай да бір мүшелерінің рак ауруымен қатар байқалатын моногендік аурулардың бірқатар түрі бар.2)Ұрпақтан ұрпаққа тұрақты тұқым қуалайтын:жатырдың,аналық бездің,сүт бездерінің,өңеш рагы.3)Барлық ісік жасушалары хрормосомалар мен ДНҚ-ның құрылысындағы және атқаратын қызметіндегі анық байқалатын бұзылулармен сипатталады.
Ісіктің аса маңызды суперессор гендерінің бірі р-53 белогының гені.Оның көптеген аллельдерң бар.Адамның барлық ісік ауруларының жартысына жуығы осы р-53 генінің мутантты аллельдерінің бақылауымен қалыптасады.Мутация нәтижесінде р-53 генінің екі аллельінің активсізденуі қауіпсіз аденоманы қатерлі карциномаға айналдырады.Ісіктің нақты бір жерде орналасуын бақылайтын геннің мутацияға ұшырауы,оның түрлі мүшелерге таралып,метастазалануына себеп болады
Митоздық циклдың реттелуіндегі циклин және циклинге тәуелді киназалар рөлінің сызбасын сызу және түсіндіру
Жасушалық циклда арнайы протин-киназалар-циклин тәуелді киназалар –ЦТКшешуші рөл атқарады. АКтивті киназалар циклин-циклин тәуелді киназалар кешені түрінде болады. Бұл кешенде циклин активтендіруші болса ЦТК-лар катализаторлар қызметін атқарады. Митотикалық циклдың инициациясы циклин-Д-ЦТК-4және циклин –Д-ЦТК-6кешенің әсеріне байланысты жүреді. Олар посмитотикалық кезеңінің алғашқы сатысында қызмет атқарып жасушаның G1 кезеңінен S-кезеңіне өтуге жағдай туғызады S- кезеңде бірінен соң бірі циклин-А-ЦТК- 2кешені және циклин-В-ЦТК-2кешендері қызмет атқарып ДНҚ репликациясына қатысатын басқа арнайы белоктарға әсерін тигізеді.G2- кезеңінде реттеуші фактор ретінде циклин-в-ЦТК кешені қызмет атқарады. Бұл кезең жасушаныңG2- кезеңінен митозға ауысу процессін қамтамасыз етіп митоздық бөлінудің жүруін бақлайды. Оны стимулдаушы фактор деп атайды.
Митоздық циклдың бақлануындағы тексеру нүктелерінің сызбасын сызу және түсіндіру
Митоздық цикл-жасушаның бөлінуге даярлануы бөлінуі кезеңдерінде жүретін сатылы және бір-бірімен тығыз байланысты процестердің жиынтығы Бұл цикл 2 кезеңнен тұрады:
1)интерфаза бұл кезеңде жасушаның белсенді өсуіне қажет ақуыз РНҚ және басқа заттар синтезделеді және ДНҚ репликациясы жүреді.
2) жасушаның бөлінуі( митоз)
Кезеңдері:
1)Интерфазаның бастапқы кезеңінде интерфазалық жасушаның құрылымдық ерекшіліктері қалпына келтіреді жасушада қарқынды түрде биосинтез процессі жүреді. Ең ұзақ кезең ұзақтығы 10сағ-тан бірнеше тәулікке созылады.
2)Синтезделу кезеңі(S) жасушаның тұқым қуалау материалы ДНҚмолекуласының екі еселенуімен сиппатталады ДНҚ-тізбектері бір-бірінен ажырап әрқайсысының жанынан комплементарлы жаңа тізбектер синтезеледі Тұқым қуалау матириалы екі есе артады
3)Синтезден кейінгі(G2) –ДНҚ-синтезі тоқталып қарқынды түрде энергия қоры жинақтала бастаиды. Ең қысқа кезең ұзақтығы 3-4сағат
Молекулалық генетикалық әдіс – ДНҚ молекуласын амплификациялау және секвинирлеу әдісінің сызбасын сызу және маңызын түсіндіру.
Молекулалық генетикалық әдістерді жасушаның генетикалық материалының құрылысы мен қыметін ДНҚ молекуласының деңгейінде зерттеу үшін қолданады.
1)ДНҚ фрагменттерін амплификациялау (көптеген көшірмелер алу)
2)Секвенирлеу- тұтас ДНҚ молекуласындағы және оның фрагменттеріндегі нуклеотидтердің орналасу ретін анықтау.
ДНҚ-полимеразалардың зерттелетін ДНҚ фрагментінің амплификациясына қатынасады.Амплификация түрлерінің бірі-ПТР әдісін пайдалану арқылы ДНҚ-ның арнайы амплификациясы іске асырылады.Ол үшін ДНҚ-полимераза ферменті екі праймердің (қоздырғыштың) қатынасуымен ДНҚ молекуласының өзара комплементарлы тізбектерін таңдамалы түрде синтездейді.Амплификацияланатын фрагменттердің ұзындығы праймерлердің арақашықтығымен анықталады.Амплификациялауға қажетті нуклеотидтік жүйелері бар кез-келген ДНҚ үлгілерін матрица ретінде пайдаланып,зерттелетін ДНҚ фрагментінің жүздеген миллион көшірмелерін алуға болады.Полимеразалық тізбектік реакция жүргізуге қажетті жағдайлар:1)ұзындығы 100-ден 35000 жұп нуклеотидтер дейінгі матрицалық ДНҚ-нысаналар 2)екі,жасанды синтезделген,ұзындығы 15-30 ж.н. праймерлер-олигоуклеотидтік жүйелер 3)Жоғары температураға тұрақты,өзінің активтілігін 94С та не одан жоғары температурада сақтай алатын ДНҚ полимеразалар 4)дезосирибонуклеотидтің барлық төрт түрі-адениндік,тиминдік,цитозиндік,гуаниндік нуклеотидтер
Секвенирлеу-ДНҚ фрагменттеріндегі нуклеотидтердің орналасу ретін анықтау әдісі.Ол үшін ұзындықтары бар болғаны бір азотты негіз бойынша түрліше болып ажыратылатын ДНҚ-ның комплементарлық молекулаларының жүйелері алынады. Екі әдісі бар:1)Максам-Гилберт әдісі-бір азотты негіз бойнша ДНҚ-ны химиялық жолмен ажыратуға негізделген.
2)Сангер әдісі не дидезокси – әдісі – өте қарапайым және сенімді әдіс болғандықтан тәжірибелік жұмыстарды жиі қолданады.Секвенирлеу жүргізу үшін мыналар қажет:1)секвенирлеуші праймер (бастапқы ДНҚ молекуласының нақты учаскесіне комплементарлы келетін жасанды синтезделген нуклеотидтер жүйесі).2)4дезоксинуклеотидтердің жиынтығы бар төрт пробирка,әр пробиркадағы 4 d АТФ;d ЦТФ;d ГТФ;d ТТФ дезоксинуклеотидтердің біреуі сол пробиркаға қосылатын дидезоксинуклеотидтің түріне сәйкес изотопты таңбаланған болады.
Секвенирлеу ДНҚ-ның зерттелетін фрагментінің құрамындағы барлық октаннуклеотидтерді анықтайды.Олардың орналасу реті арнайы компьютерлік бағдарламаның көмегімен айқындалады.
Оогенез кезеңдерінің сызбасын сызу және түсіндіру.
Гаметогенез


Оогенез Сперматогенез
Оогенез-аналық жыныс безінде өтеді.
3кезеңнен тұрады:
1кезең-Көбею кезеңі –овогониялар түзіледі.
2кезең-Өсу кезеңі-овоциттер қалыптасады.
3кезең-Пісіп жетілу кезеңі-мейоз жүреді.
Оогенездің пісіп жетілу аймағында мейоздың әр бөлінуі сайын түзілген жас жасушаларда цитоплазма біркелкі бөлінбейді. Нәтижесінде бір ірі жұмыртқа жасушасы және 3цитоплазмасы жоқ бағыттаушы денелер түзіледі.Оогенезде бір ғана жұмыртқа жасушасы пайда болады
Онтогенздің кезеңдерінің сызбасын сызу және адам онтогенезінің қатерлі кезеңдерінің маңызын түсіндіру.
Онтогенез-дараның жеке дамуы немесе дараның зигота түзілуімен басталып тіршілігін жойғанға
дейінгі дамуы. Кезеңдері:
1.прогенез (преэмбриональдық кезең);
2.антенатальды (туылғанға дейінгі)кезең;
Сатылары:
1)бөлшектену 1-4күн; митоз жолымен синхронды емес жүреді,бласт.саны өскенмен мөлшері өспейді. Оплазмалық сегригация
2)бластрула 5-8күн; көп жасушалы бірқабатты ұрық. Позициялық ақпарат
3)гаструла 9-14күн; 1не3 қабатты ұрық. Бұл кезде ұрық жапырақшаларының түзілуі жасушалардың таңдамалы сұрыпталуы нәтижесінде жүзеге асады. Детерминация-эмбриональды бастамалардың бағытының даму жолын анықтау.
4)нейрула, морфогенез 15-17күн;(гисто- және органогенез). Дефференциация-бұл үрдістің нәтижесінде жасуша арнайыланады, яғни белгілі бір химиялық, морфологиялық және функционалдық ерекшелікке ие болады. Эмбриональды индукция
3.постнатальды (туылғанан кейінгі) кезең;
Онтогенездік дамуды қамтамасыз ететін гендердің иерархиялық жүйесінің сызбасын сызу және түсіндіру.
Онтогенез – дараның жеке дамуы немесе дараның зигота түзілумен басталып тіршілігін жойғанға дейінгі дамуы.Онтогенез ұрықтыну кезінде ата-аналарының жыныс жасушаларынан алынған тұқым қуалайтын ақпараттың негізінде жүзеге асады.Онтогенездік дамуды екі кезеңге бөледі:
Туылғанға дейін(антенатальді),
Туылудан кейін (постнатальді) кезең.
Онтогенездің ерекше кезеңі – предэмбриональды немесе гаметогенез кезеңі бөледі.
Гаметогенез-жұмыртқа жасушасының және сперматозоидтардың түзілу процесі.
Онтогенездік дамудың ерте кезеңіндегі жасушалық механизмдердің сызбасын сызу және түсіндіру.
Онтогенез дараның жеке дамуы немесе дараның зигота түзілуімен басталып тіршілігін жойғанға дейінгі дамуы.Онтогенез механизміне мына процестер жатады:пролиферация,миграция,жасушаның сұрыпталуы,апоптоз.
Пролиферация(көбею) біржасушалы зиготаның бөлінуі арқылы көпжасушалы ұрық дамиды.Бөліну ағзаның өсуін,қалыпты дамуын қамтамасыз етеді.Мысалы:мутациялардың әсерінен жасушалардың көбеюін бақылаудың жойылуы қатерлі ісіктердің пайда болуына әкеп соқтырады.
Жасуша миграциясы не орын ауыстыруы онтогенез процесінде үлкен рөлі бар және гаструляцмя,морфогенез үдерістерін қамтамасыз етеді.Оның бұзылуы туа біткен даму ақаулықтарына,гетеротопияға алып келеді.Мысалы:нейропластар миграциясының бұзылуы,микро-макрогирия,полигирия,агирияға әкеледі.
Жасушалардың сұрыпталуы олардың таңдамалы адгизияға қалыпты дамуды қамтамасыз етіп, морфогенезде маңызды рөл атқарады және генетикалық бақылау мен қоршаған жасушалар әсерінің бақылауында болады.Жасушалар эмбриогенез үдерісінде тек қана белсенді көшіп қана қоймай,бірін бірі танып тек белгілі бір жасушалармен пласттар түзеді.Бұл үдерістер гаструляция кезеңіне тән және жасушалардың таңдамалы сұрыпталуы деп аталады.Ісік жасушаларының метостазға қабілеттілігі таңдамалы адгезияның бұзылуына байланысты.
Жасушалардың жойылуы апоптоз немесе некрозға бөлінеді.Апоптоз қалыпты физиологиялық жағдайларға тән,жасушаның бағдарламаланған өлімі.Онтогенезде жүреті басқа да жасушалық үдерістер сияқты,апоптоз табиғи,эволюция жолында қалыптасқан және генетикалық бақыланатын үдеріс болып табылады.Апоптоз генетикалық факторлармен және жасушааралық өзара әсермен бақыланады.Қалыпты онтогенезді к даму барысында апоптозға ұшырайтын жасушаларда олардың тірщілігін қамтамасыз ететән гендердің белсенділігі уақытпен щектелген болады.Басқаша айтсақ,мұндай жасушалар тіршілігінің белгілі кезеңінде олардың генетикалық бағдарламаланған өлімі.Адамда апоптоз үдерісінің бұзылуы (мутагендік не тератогендік) синдактилия (қол,аяқ саусақтарының бітісуі),ішектің бітелуі және дамудық туа біткен ақаулықтарына алып келеді.Соңғы кездері жасушадық бөлінуді тоқтату үдерісін бақылайтын және апоптоз механизмін іске қосатын ген ашылды.Бұл геннің,р53 мутациясы жасушалардың қатерлі ісікке айналуына алып келеді.Мыс:Аналық дараларда вольф каналының дегенерациясы,аталық дараларда мюллер каналының дегенерациясы.
Онтогенездік дамудың ерте кезеңінде жұмыртқа жасушасының полярлығы,ооплазмалық сегрегация,позициялық ақпарат,детерминация және дифференциацияның мәні мен маңызын түсіндіру және сызбасын сызу.
Жұмыртқа жасушасында сарыуыздың орналасуының маңызы зор,өйткені болашақта ұрықтың кеңістіктік құрылымын анықтайды.Сарыуыздың орналасу сипатына байланысты жұмыртқа жасушасының полярлығы:бұл ядроның эксцентрлік орналасып,анимальды полюске қарай ығысуын және цитоплазмалық қосындылардың орналасу ерекшеліктерін көрсетеді.
Жұмыртқа жасушасының цитоплазмасында пайда болған жергілікті айырмашылық,оның ішкі сапасының әртүрлі болуына алып келеді,оны ооплазмалық сегрегация деп атайды.ОС ұрықтанудан кейін күшейеді де,ұрықтың бастапқы жіктелуіне негіз болады.
Жасушаның позициялық ақпаратына сәйкес мүшенің бастамасы өзінің орналасуын координаттық жүйе бойынша бағалап,содан кейін сол жағдайға байланысты жіктеледі.Жасушаның орналасуы ұрықтың ұзына бойы осіндегі белгілі градиентпен оргаласқан кейбір заттардың концентрациясымен анықталады.
Детерминация – (анықтау,шектеу) морфогенездің алғашқы кезеңдерінде дамып келе жатқан ағза бөлімдері арасында сапалық айыпмашылықтардың пайда болуы.Детерминацияның негізінде кейбір гендердің активтелінуі және м-РНҚ МЕН ақуыздың синтезі жатыр.Детерминация және эмбриональдық реттеу қарама-қарсы қасиеттер.Детерминация бүтіннен бөліктерге қарай жүреді-бірінші ұрықтың бүтін бастамасында детерминация жүреді,ал оның жеке элементтерінің тағдыры әлі белгісіз.Қалыпты даму кезінде жауапты материалда индуктордың әсерінен алғашқыда тұрақсыз (лабильды)д-я,қайтымсыз д-я жүреді.
Жасушалардың дифференциациялануы - арнайы жасуша,мүшелер мен ұлпалардың қалыптасыуына әкелетін,дараның даму барысында біртекті жасушалар мен ұлпалардың арасында айырмашылықтардың пайда болуы және олардың өзгерулері.Негізгі факторлары-алғашқы эмбриональды жасушалардың цитоплазмасының ерекщелігі және көршілес жасушалардың арнайы әсері – индукция.Этаптары:1)жасушалардың дифференциациялануының 1-ші себебі жұмыртқа жасуша цитоплазмасының химиялық әртүтлілігі б.т.2)ол ұрықтанудан кейін күшейеді:сперматозоид жұмыртқа жасушасында енген кезде цитоплазма компоненттерінің қайта орналастыруы(ооплазматикалық сегрегация)3)жұмыртқа жасушасы цитоплазманың химиялық әртүрлілігі бластомерлердің цитоплазмасының химиялық әртүрлілігі ауысады.4)әр түрлі бластомерлерде әр түрлі индукторлар болады.5)әр түрлі ұлпалардан әр түрлі мүшелер түзіледі
Протоонкогеннің онкогенге айналу механизмдерінің сызбасын сызу және түсіндіру.
Протоонкогендер деп онкогенге ұқсас қалыпты не ізашар гендерді айтады.Протоокогендер кәдімгі «қалыпты» гендер.Олар басқа гендермен бірге жасушалардың бөлінуін,өсуін,өзара әрекеттесулерін реттейді.Бұл гендер «өсу факторлары» деп аталатын белоктың синтезін бақылайды.
Протоонокегенннің онкогенге айналуы мына жолдармен іске асырылады:
1.протоонкогенге жаңа транскрипцияның жалғануы;
2.протоонкогеннің онкогендік активтілікке дейінгі амплификациясы(көшірмелерлі көбейту);
3.Иелік-жасушаның ДНҚ молекуласының кейбір нуклеотидтер жүйесінің қосылып,промотрдың әрекетін күшейту;
4.транслокация нәтижесінде протоонкогеннің иммуноглобиннің локусына жалғануы;
5.протоонкогеннің мутациялары;
Хромосомалардың транслокациясы нәтижесінде протоонкогенге жаңа тарскрипциялық промотордың жалғануы протоонкогеннің үздіксіз жұмыс істеп,белоктың тоқтаусыз синтезделуіне,яғни оның онкогенге айналуына себеп болады.Олай болса,протоонкогеннің онкогенге айналуына себеп болатын негізгі және жалпы механизмдерінің бірі протоонкогендер активтілігінің қалыпты реттелуі мен бақылауының бұзылуы.Протоонкоген К-ras-тың онкогенге айналуы ісіктің өсіп,ұлғаюына алып келеді.Ісіктің аса маңызды суперессор гендерінің бірі р-53 белогының гені.Оның көптеген аллельдерң бар.Адамның барлық ісік ауруларының жартысына жуығы осы р-53 генінің мутантты аллельдерінің бақылауымен қалыптасады.Мутация нәтижесінде р-53 генінің екі аллельінің активсізденуі қауіпсіз аденоманы қатерлі карциномаға айналдырады.Ісіктің нақты бір жерде орналасуын бақылайтын геннің мутацияға ұшырауы,оның түрлі мүшелерге таралып,метастазалануына себеп болады.
Рекомбинативті өзгергіштіктің пайда болу механизмдерінің сызбасын сызу және оның генетикалық маңызын түсіндіру.
Өзгергіштік-бұл тірі ағзалардың орта факторларының әсерінен жаңа белгілерге ие болу қасиеті.Өзгергіштік фенотиптік (тұқым қуаламайтын) және генотиптік (тұқым қуалайтын)болып бөлінеді.Рекомбинативтік өзгергіштік генотиптік өзгергіштіктің бір түрі,бұл жағдайда бір түрге жататын ағзалар бір бірінен генотиптегі гендердің басқаша топтасуының нәтижесінде ажыратылады.Бұл ата анасына тән емес жаңа белгілердің пайда болуына алып келеді.Мысалы,ата-аналарының қан топтары 2 мен 3 болса,баласының қан тобы 4 болуы мүмкін.
Рекомбинативтік өзгергіштіктің механизмдері:
1.Гаметогенез кезінде мейоздың анафазасында әр гомологты жұптарындағы аталық және аналық хромосомалардың тәуелсіз ажырауы;
2.Кроссинговердің нәтижесінде генетикалық материалдың рекомбинациясы;
3.Ұрықтану кезінде гаметалардың кездейсоқ қосылуы;
Рекомбинативтік өзгергіштіктің биологиялық маңызы өте зор,себебі ол бір түрге жататын даралардың генетикалық әртүрлілігін,оның өкілдерінің тірі қалуын қамтамасыз етеді.
Сызықты ДНҚ молекуласының репликацияланбау себептерін және ұштарының ұзару механизмдерінің сызбасын сызу
Репликация процесінде ДНҚ-ның жаңа тізбектері синтезделместен бұрын алдымен РНҚ-праймердің синтезделіп қызметін атқарып соңынан оның шығарылып тасталады .Эукариоттар репликациясының ерекшілігі осы синтезделген РНҚ-праймер бөлініп кеткен соң әр репликация аяқталғанда түзілген ДНҚ-ның жаңа тізбектері бастапқы матрициялық тізбектен сәл қысқалау келеді. Жаңадан синтезделген тізбектің жетіспейтін бөлігінің ұзындығы РНҚ-праймердің мөлшеріне 10-нуклеотидке сәйкес болады. Бастапқы матрициялық тізбектің 3-ұшынан жаңа тізбектің комплементарлы5-ұшы қысқа болып аяқталады.
Теломералардың болуы ДНҚ-ның тұқым қуалау ақпараты жазылған учаскелерінің репликацияланбай қалуынан сақтайды. Жасушаның әр бөлінуі сайын түзілген жас жасушалардағы хромосомалардың теломералық учаскелерінің қысқаруы ең соңында олардың толық жойылып бірте-бірте ақпараты бар гендердің толық репликацияланбай қалуына себеп болады .
Теломераза репликация аяқталардан бұрын ДНҚ-ның жаңа синтезделген тізбектерінің толық репликацияланбаған 5-қштарын қысқалау келген қайталанатын нуклеотидтер жүйесімен немесе теломералармен жалғап ұзартады.
Сперматогенез кезеңдерінің сызбасын сызу және түсіндіру.
Cперматогенез-аталық бездерінің түтікшелерінде жүреді бірнеше сатылардан тұрады. Ағза жыныстық жетілген кезінен бастап түтікшелердің сыртқы қабатындағы диплоидты спермотогониялар қарқынды түрде митоздық бөлінуін бастайды. Бұл-көбею аймағы. Сперматогониялардың бір бөлігі келесі өсу аймағына өтеді. Бұл кезеңде цитоплазма көбейіп олардан бірінші реттегі сперматоциттер түзіледі. Келесі аймақ-пісіп жетілу аймағы Мұнда -мейоз жүреді. Сперматогенездің ерекшелігі қалыптасу аймағы. Бұл аймақта сперматидтердің ядросы тығыздалып акросома аппараты қалыптасады және цитоплазма мөлшері азаяды. Сперматидтер кішірейеді құйрығы пайда болып нағыз сперматозоидтарға айналады
Тірі жүйелердегі тұқым қуалау ақпаратының тасымалдану типтерін және бағыттарын сызу және түсіндіру.
Тұқым қуалау ақпараты-ДНҚ-да тізбектесе орналасқан нуклеотидтер ретімен жазылған.Ген-тұқымның бірлігі ретінде бір полипептидтік тізбек синтезделетін ,бақыланатын ДНҚ учаскесі.
Тұқым қуалау ақпаратының берілу типтері.
1. Жалпы берілу: кез-келген жасушаларда беріледі.
2.Арнайы берілу:кейбір ерекше жағдайдағы арнайы жасушада жұреді.
Тұқым қуалау ақпаратының жалпы берілу типі:
1)ДНҚ-ДНҚ (репликация)
2)ДНҚ-а-РНҚ( транскрипция)
3)а-РНҚ-ақуыз-(трансляция)
Тұқым қуалау ақпаратының арнайы берілу типі:
1)РНҚ-РНҚ (РНҚ репликациясы)
2)РНҚ-ДНҚ (кері тарнскрипция)
3)ДНҚ-белок (ДНҚ трансляциясы)
Тұқым қуалау ақпаратының жүзеге асырылу кезеңдері:
1)Репликация.
2)Транскрипция
3)Трансляция
Транскрипция үрдісінің кезеңдерінің сызбасын сызу және транскрипцияға қатысатын ферменттерді сипаттау.
Генетикалық ақпараттың ДНҚ-дан а-РНҚ-ға көшіріліп жазылу процесін транскрипция д.ат.
Транскрипция процесі ядрода арнайы фермент РНҚ-полимеразаның көмегімен ДНҚ және синтезделетін а-РНҚ нуклеотидтердің комплементарлы жұптасуы жолымен іске асырылады.А-РНҚ-ның тізбегінің синтезделуі 51→31бағытында жүреді,яғни жаңа нуклеотидтер синтезделетін а-РНҚ тізбегінің тек 31-ОН ұшына жалғанады.Транскрипция тұтас бір хромосоманың бойында емес,тек ДНҚ-ның бір тізбегінің жеке учаскелерінде жүреді.ДНҚ молекуласының екі тізбегі бір-бірінен қызметтері бойынша ажыратылады.Тізбектердің біреуі мағыналы немесе кодтаушы ,екіншісі матрицалық тізбек деп аталады.
Прокариоттардағы транскрипция.
Прокариоттарда транскрипция 3 кезеңнен тұрады:1.Инициация. 2.Элонгация. 3.Терминация.

Транскрипция процесі 51 →31 бағытында жүреді.
Матрицалық ДНҚ-ның транскрипция жүретін учаскесі кодтаушы учаске д.ат.
Промоторда орналасқан арнайы нуклеотидтер жүйесі старттық нүкте болып табылады,оны -35жүйелік д.ат.9 нуклеотидтен тұратын консенсустық жүйе д.ат.
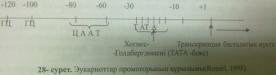
Прокориоттар промоторы 20-200 нуклеотидтен тұрады,оның құрамында 7 нуклеотидтен құралған арнайы жүйе ТАТАААТ бар,оны Прибнов-боксы д.ат.
Эукариоттар транскрипциясы 3 кезеңнен тұрады.1.Инициация.2.Элонгация.3.Терминация.
Эукариоттар транскрипциясында РНҚ-полимеразаның 3 түрі қатысады:РНҚ-полимераза І,ІІ,ІІІ салмағы 500 000 дальтон Эукариоттар генінің промоторында 7 нуклеотидтен тұратын : ТАТАААТ консенсустық жүйелік болады,оны Хогнесс-боксы немесе ТАТА –бокс дейді. Транскрипциялық факторлар –транскрипцияны бастауға қажетті белоктар.
Хромосомалық мутациялар хромосома құрылысының өзгеруімен. Олар хромосома ішілік, хромосома аралық болып бөлінеді.
-
хромосома ішілікке:
Делеция(жетіспеушілік)- хромосома бөлігінің түсіп қалуы.
Дупликация- хромососма бөлігінің екі еселенуі.
Инверсия- хромосома бөлігінің үзіліп 1800 бұрылып сол хромосомадағы орнына қайта жалғануы; перицентрлік-центромера аймағында, парацентрлік-хромосоманың бір иығында жүріп, центромерадан алшақ байқалады;
-
хромосома аралыққа:
Транслокация- гомологтық емес хромосомалардың учаскелерімен алмасуы; Оның бірнеше түрі бар: Реципрокты- гомологтық емес хромосомалар бір-бірімен бөліктерімен алмасады;
Реципрокты емес- бір хромосома екінші гомологты емес хромосоманың бөлігін жалғастырып өз көлемін ұзартады.
Робертсондық транслокация- гомологты емес екі акроцентрлік хромосомалар ұзын иықтарымен центромерлік аймақтары арқылыжалғанады.
