Файл: Я. А. Анатомия центральной нервной системы для психологов спб. Питер, 1е издание, 2009 год, 128 стр., формат 14x21 см (60х9016), Мягкая обложка, isbn 9785911802714 Серия Учебное пособие.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.12.2023
Просмотров: 195
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Глава 11
Кора больших полушарий
11.1. Общий план строения коры больших полушарий
Кора больших полушарий представляет собой слой серого вещества, покрывающий поверхность полушарий, толщиной от 3 до 4 мм, местами — до 5 мм. Большая часть коры (2/3) залегает в глубине борозд и не видна снаружи. Благодаря такой организации мозга в процессе эволюции была получена возможность значительно увеличить площадь коры при ограниченном объеме черепа. Общее число нейронов коры мозга человека оценивается в 10-15 млрд.
Кора большого мозга является наиболее высоко дифференцированным отделом нервной системы, однако она неоднородна. В соответствии с филогенезом различают древнюю, старую и новую кору. Древняя кора {paleocortex) включает в себя неструктурированную кору вокруг переднего продырявленного вещества: околоконечную извилину, подмозолистое поле (расположено на внутренней стороне полушарий под коленом и клювом мозолистого тела). Старая кора (archicortex) двух-трехслойна, расположена в гиппокампе и зубчатой извилине. Новая кора (neocortex) составляет 96% от всей поверхности полушарий. По морфологическим особенностям в ней выделяют шесть основных слоев, однако в различных областях коры количество слоев варьирует (рис. 11.1).
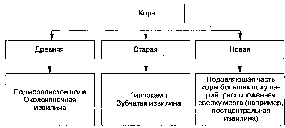
Рис. 11.1. Схема классификации коры
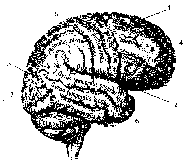
Рис. 11.2. Латеральная поверхность коры больших полушарий [9]:
1 — центральная борозда; 2 — боковая борозда; 3 — теменно-затылочная
борозда; 4 — лобная доля; 5 — теменная доля; 6 — височная доля;
7 — затылочная доля
Кора полушарий покрыта бороздами и извилинами. Различают самые глубокие и постоянные с точки зрения изменчивости первичные борозды, которые делят полушария на доли. Боковая борозда (силъвиева) отделяет лобную долю от височной, центральная борозда (ролапдова) — лобную от теменной. Темен-но-затылочная борозда располагается на медиальной поверхности полушария и разделяет теменную и затылочную доли (на верхнелатеральной поверхности явная граница между этими долями отсутствует) (рис. 11.2). На медиальной поверхности располагается поясная борозда, переходящая в гиппокампову борозду. Они ограничивают обонятельный мозг от остальных долей.
Вторичные борозды менее глубокие, они делят доли на извилины и тоже достаточно постоянны. Третичные (безымянные) борозды придают извилинам индивидуальную форму и существенно увеличивают площадь их коры. Они наиболее изменчивы.
11.2. Макроскопическое строение коры
11.2.1 Верхнелатеральная поверхность полушарий
Лобная доля. В заднем отделе наружной поверхности этой доли проходит прецентральная борозда. От нее в продольном направлении проходят две борозды: верхняя и нижняя лобные. Благодаря этому лобная доля разделяется на четыре извилины: прецент-ральную, верхнюю лобную, среднюю лобную и нижнюю лобную.
Теменная доля. На ней располагается постцентральная борозда, сливающаяся обычно с межтеменной бороздой, В зависимости от расположения этих борозд теменная доля разделяется на три извилины: постцентральную извилину, верхнюю теменную дольку и нижнюю теменную дольку.
Височная доля. Латеральная поверхность этой доли имеет три продольные извилины, отграниченные друг от друга верхней pi нижней височными бороздами. Этими извилинами являются верхняя, средняя и нижняя височные извилины.
Затылочная доля. Борозды латеральной поверхности этой доли весьма изменчивы и выделяют только поперечную затылочную борозду.
Островок. Эта долька имеет форму треугольника. Поверхность островка покрыта короткими вариабельными извилинами.
11.2.2. Нижняя поверхность полушарий
В лобной доле проходит обонятельная борозда. В височной доле видны две борозды: затылочно-височная, проходящая в направлении от затылочного полюса к височному и ограничивающая латеральную затылочно-височную извилину, и идущая параллельно ей коллатеральная борозда. Между означенными бороздами располагается медиальная затылочно-височная извилина. Медиально от коллатеральной борозды расположены две извилины: язычок и парагиппокампальная извилина. Последняя, примыкая к стволу мозга, находится уже на медиальной поверхности полушария.
11.2.3. Медиальная поверхность полушарий
На медиальной поверхности мозга выделяют борозду мозолистого тела, идущую непосредственно над мозолистым телом и продолжающуюся своим задним концом в глубокую гиппокам-пальную борозду. Выше расположена поясная борозда, которая ограничивает поясную извилину и продолжается в парагиппо-кампальную извилину, заканчивающуюся крючком
{uncus). Эти образования тесно связаны с лимбической системой. Небольшой участок коры, ограниченный поясной и парацентральными бороздами, называется парацентральная долька. Сзади от пара-центральной дольки находится предклинье, относящееся к теменной доли. Позади предклинья лежит обособленный участок коры, относящийся к затылочной доле — клин. Клин отделяется от предклинья глубокой теменнозатылочной бороздой.
11.3. Микроскопическое строение коры
11.3.1. Цитоархитектоника
Клеточную организацию коры больших полушарий называют цитоархитектоникой. В коре больших полушарий можно выделить шесть слоев, которые различаются как по морфологиче ским особенностям, так и по функциональному предназначению (рис. 11.3).
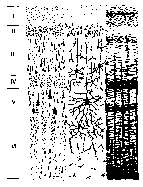
Рис. 11.3. Микроскопическое строение коры больших полушарий:
I — молекулярный слой; II — наружный зернистый слой; III — наружный
пирамидный слой; IV — внутренний зернистый слой; V — внутренний
пирамидный слой; VI — ганглионарный слой.
Первая колонка — тела клеток коры, прокрашенные без отростков; вторая
колонка — нейроны коры с отростками; третья колонка — нервные сплетения
коры больших полушарий
Морфологически различают: гранулярную кору (в ней наиболее выражены II и IV слои клеток округлой формы) и агрануляр-ную кору (в ней наиболее выражены III, V и VI слои, богатые пирамидными нейронами). К гранулярной коре относится сенсорная кора — зрительная, слуховая и пр., а к агранулярной — двигательная и ассоциативная кора. Дифференцировка слоев происходит с 6 по 8 месяц внутриутробного развития. Каждый слой коры отличается по своим морфологическим особенностям (табл. 11.1).
Таблица 11.1 Слои коры
Клетки различных слоев коры объединены в так называемые «модули», являющиеся структурно-функциональными единицами коры. Как правило, это группы нейронов из 100-1000 клеток, которые выполняют определенные функции, «обрабатывая» определенный сигнал.
Обычно эти группы клеток напоминают вертикальные колонки. Впервые «колонки» в коре обнаружил В. Маунткасл, а в нашей стране исследованием «корковых модулей» занимался А. Б. Коган и его последователи из ростовской нейрофизиологической школы.
Особая роль этих кортикальных колонок в «ассоциативных» полях, а также во всей коре, вероятно, связана с их способностью к установлению ассоциаций, т. е. образованию временных связей, с другими колонками из других участков мозга.
11.3.2.. Миелоархитектоника
Рис. 11.4. Миелоархитектоника коры больших полушарий
Так как кора больших полушарий является высшим информационным центром нервной системы человека, то в коре имеет место интенсивный обмен нервными импульсами, как между отдельными ее участками, так и с другими отделами центральной нервной системы. Обмен информацией осуществляется по волокнам (белое вещество), соответственно, количество нервных волокон в больших полушариях очень велико. Сложно организованную систему волокон коры больших полушарий называют миелоархитектоникой. При этом всю совокупность нервных волокон можно разделить на несколько групп (рис. 11.4).
Основные пучки внутрикорковых волокон располагаются в первом слое (тангенциальные волокна), во втором слое (полоска Бехтерева), в четвертом и пятом слоях (соответственно наружная и внутренняя полоски Байарже). Эти нервные пучки отражают распространение афферентных волокон и коллате-ралей аксонов пирамидных клеток.
11. 4 Функциональное значение отдельных зон коры
Два полушария, составляющие передний мозг, действуют согласованно. Правое полушарие контролирует сенсорные и двигательные функции левой половиргы тела, а левое осуществляет аналогичный контроль над правой половиной. Связь между зрительными зонами левого и правого полушарий в норме осуществляется через мозолистое тело.
Функционально различают три разновидности коры: чувствительная
(сенсорная), двигательная (моторная) и ассоциативная (рис. 11.5).
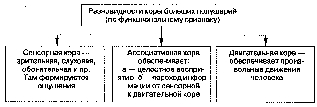
Рис. 11.5. Разновидности коры больших полушарий по функциональному признаку
Как понятно из названия, сенсорная кора ответственна за обработку информации, поступающей от органов чувств. Именно в ней располагаются корковые отделы анализаторов человека: в затылочной области — зрительного, в височной — слухового, в теменной — кожного и т. д.
В двигательной коре располагаются первые нейроны, управляющие работой произвольных мышц человека. Каждой мышечной группе соответствует определенная область двигательной коры (рис. 11.6).
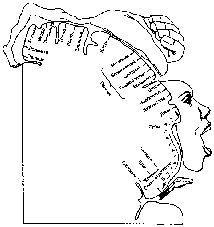
Рис. 11.6. Представительство различных мышечных групп в коре больших полушарий человека («человечек Пенфилда») [1]
Подавляющая часть корковой площади занята именно ассоциативной корой. Как полагает большинство ученых, именно там образуются ассоциативные связи между специализированными областями и интегрируется приходящая из них информация. Кроме того, здесь, как полагают, текущая информация объединяется с эмоциями и воспоминаниями, что позволяет людям думать, решать, составлять планы [1,4]. Например, считается, что ассоциативные поля теменной доли объединяют информацию, приходящую от соматосенсорной коры (сообщения от кожи, мышц, сухожилий и суставов относительно положения тела и его движений) — со зрительной и слуховой информацией, поступающей из зрительной и слуховой коры затылочной и височной долей. Как пишет Флойд Блум и его соавторы, «эта объединенная информация помогает нам иметь точное представление о собственном теле во время передвижений в окружающем пространстве. Слияние сенсорных данных с информацией, извлекаемой из кладовых нашей памяти, позволяет нам осмысленно интерпретировать специфические зрительные сигналы, звуки и тактильные ощущения. Когда что-то движущееся и пушистое коснется вашей руки, вы будете по-разному реагировать на это в зависимости от того, услышите ли вы одновременно с этим мурлыканье вашей кошки или рев медведя» [1; с. 175]. После обработки информации в ассоциативной коре интегрированная картина события передается лобной коре. Благодаря обширным двусторонним связям с лимбической системой к этой картине добавляется эмоциональный оттенок, а также информация, взятая из памяти. Другие нервные связи тоже доставляют информацию, которая позволяет лобной коре оценить текущие требования организма и окружающей среды и выбрать среди них первоочередные — решить, что лучше, а что хуже для организма в данной ситуации. Лобная кора, по-видимому, ответственна и за выбор целей, которые мы ставим перед собой на будущее, а также за нашу оценку различных обстоятельств в связи с этими целями [1].
Кора больших полушарий
11.1. Общий план строения коры больших полушарий
Кора больших полушарий представляет собой слой серого вещества, покрывающий поверхность полушарий, толщиной от 3 до 4 мм, местами — до 5 мм. Большая часть коры (2/3) залегает в глубине борозд и не видна снаружи. Благодаря такой организации мозга в процессе эволюции была получена возможность значительно увеличить площадь коры при ограниченном объеме черепа. Общее число нейронов коры мозга человека оценивается в 10-15 млрд.
Кора большого мозга является наиболее высоко дифференцированным отделом нервной системы, однако она неоднородна. В соответствии с филогенезом различают древнюю, старую и новую кору. Древняя кора {paleocortex) включает в себя неструктурированную кору вокруг переднего продырявленного вещества: околоконечную извилину, подмозолистое поле (расположено на внутренней стороне полушарий под коленом и клювом мозолистого тела). Старая кора (archicortex) двух-трехслойна, расположена в гиппокампе и зубчатой извилине. Новая кора (neocortex) составляет 96% от всей поверхности полушарий. По морфологическим особенностям в ней выделяют шесть основных слоев, однако в различных областях коры количество слоев варьирует (рис. 11.1).
Рис. 11.1. Схема классификации коры
Рис. 11.2. Латеральная поверхность коры больших полушарий [9]:
1 — центральная борозда; 2 — боковая борозда; 3 — теменно-затылочная
борозда; 4 — лобная доля; 5 — теменная доля; 6 — височная доля;
7 — затылочная доля
Кора полушарий покрыта бороздами и извилинами. Различают самые глубокие и постоянные с точки зрения изменчивости первичные борозды, которые делят полушария на доли. Боковая борозда (силъвиева) отделяет лобную долю от височной, центральная борозда (ролапдова) — лобную от теменной. Темен-но-затылочная борозда располагается на медиальной поверхности полушария и разделяет теменную и затылочную доли (на верхнелатеральной поверхности явная граница между этими долями отсутствует) (рис. 11.2). На медиальной поверхности располагается поясная борозда, переходящая в гиппокампову борозду. Они ограничивают обонятельный мозг от остальных долей.
Вторичные борозды менее глубокие, они делят доли на извилины и тоже достаточно постоянны. Третичные (безымянные) борозды придают извилинам индивидуальную форму и существенно увеличивают площадь их коры. Они наиболее изменчивы.
11.2. Макроскопическое строение коры
11.2.1 Верхнелатеральная поверхность полушарий
Лобная доля. В заднем отделе наружной поверхности этой доли проходит прецентральная борозда. От нее в продольном направлении проходят две борозды: верхняя и нижняя лобные. Благодаря этому лобная доля разделяется на четыре извилины: прецент-ральную, верхнюю лобную, среднюю лобную и нижнюю лобную.
Теменная доля. На ней располагается постцентральная борозда, сливающаяся обычно с межтеменной бороздой, В зависимости от расположения этих борозд теменная доля разделяется на три извилины: постцентральную извилину, верхнюю теменную дольку и нижнюю теменную дольку.
Височная доля. Латеральная поверхность этой доли имеет три продольные извилины, отграниченные друг от друга верхней pi нижней височными бороздами. Этими извилинами являются верхняя, средняя и нижняя височные извилины.
Затылочная доля. Борозды латеральной поверхности этой доли весьма изменчивы и выделяют только поперечную затылочную борозду.
Островок. Эта долька имеет форму треугольника. Поверхность островка покрыта короткими вариабельными извилинами.
11.2.2. Нижняя поверхность полушарий
В лобной доле проходит обонятельная борозда. В височной доле видны две борозды: затылочно-височная, проходящая в направлении от затылочного полюса к височному и ограничивающая латеральную затылочно-височную извилину, и идущая параллельно ей коллатеральная борозда. Между означенными бороздами располагается медиальная затылочно-височная извилина. Медиально от коллатеральной борозды расположены две извилины: язычок и парагиппокампальная извилина. Последняя, примыкая к стволу мозга, находится уже на медиальной поверхности полушария.
11.2.3. Медиальная поверхность полушарий
На медиальной поверхности мозга выделяют борозду мозолистого тела, идущую непосредственно над мозолистым телом и продолжающуюся своим задним концом в глубокую гиппокам-пальную борозду. Выше расположена поясная борозда, которая ограничивает поясную извилину и продолжается в парагиппо-кампальную извилину, заканчивающуюся крючком
{uncus). Эти образования тесно связаны с лимбической системой. Небольшой участок коры, ограниченный поясной и парацентральными бороздами, называется парацентральная долька. Сзади от пара-центральной дольки находится предклинье, относящееся к теменной доли. Позади предклинья лежит обособленный участок коры, относящийся к затылочной доле — клин. Клин отделяется от предклинья глубокой теменнозатылочной бороздой.
11.3. Микроскопическое строение коры
11.3.1. Цитоархитектоника
Клеточную организацию коры больших полушарий называют цитоархитектоникой. В коре больших полушарий можно выделить шесть слоев, которые различаются как по морфологиче ским особенностям, так и по функциональному предназначению (рис. 11.3).
Рис. 11.3. Микроскопическое строение коры больших полушарий:
I — молекулярный слой; II — наружный зернистый слой; III — наружный
пирамидный слой; IV — внутренний зернистый слой; V — внутренний
пирамидный слой; VI — ганглионарный слой.
Первая колонка — тела клеток коры, прокрашенные без отростков; вторая
колонка — нейроны коры с отростками; третья колонка — нервные сплетения
коры больших полушарий
Морфологически различают: гранулярную кору (в ней наиболее выражены II и IV слои клеток округлой формы) и агрануляр-ную кору (в ней наиболее выражены III, V и VI слои, богатые пирамидными нейронами). К гранулярной коре относится сенсорная кора — зрительная, слуховая и пр., а к агранулярной — двигательная и ассоциативная кора. Дифференцировка слоев происходит с 6 по 8 месяц внутриутробного развития. Каждый слой коры отличается по своим морфологическим особенностям (табл. 11.1).
Таблица 11.1 Слои коры
| Слой коры | Описание клеток и волокон |
| / — молекулярный | Содержит небольшое количество мелких ассоциативных клеток веретенообразной формы, аксоны которых идут параллельно поверхности коры, составляя тангенциальное сплетение. Этот слой содержит большое количество волокон — дендритов нижележащих слоев |
| II — наружный зернистый | Содержит округлые и звездчатые клетки, а также небольшое число пирамидных нейронов. Размеры — около 10 мкм. Аксоны могут распространяться в различных направлениях (уходить в белое вещество или подниматься к молекулярному слою, входя в состав тангенциального сплетения), а дендриты, как правило, располагаются в молекулярном слое |
| III — пирамидный | Наиболее широкий слой коры. Лучше всего развит в прецентральной извилине. Диаметр нейронов постепенно возрастает от 10 до 40 мкм сверху вниз. Главный дендрит пирамидных нейронов идет в молекулярный слой и там ветвится, боковые дендриты ветвятся в III слое, а аксон уходит вниз, образуя ассоциативное или комиссуральное волокно белого вещества |
| IV— внутренний зернистый | В так называемой гранулярной коре (например, затылочной доле) — развит хорошо, а в прецентральной извилине — неразвит. Образован мелкими клетками звездчатой формы |
| V — ганглионарный | Содержит крупные пирамиды, причем в двигательной коре — очень крупные, названные гигантскими пирамидами Беца. Их размер — до 100 мкм (120 х 80 мкм). Аксоны гигантских пирамид образуют кортикоспинальные пути спинного мозга и оканчиваются на его мотонейронах (одновременно отдавая ветви к базальным ядрам и мозжечку). Их деятельность обеспечивает произвольные движения человека |
| VI — полиморфный | Нейроны разной формы — звездчатые, пирамидные и веретенообразные. Их аксоны идут и вверх, и вниз и образуют ассоциативные и проекционные пути, переходящие в белое вещество головного мозга |
Клетки различных слоев коры объединены в так называемые «модули», являющиеся структурно-функциональными единицами коры. Как правило, это группы нейронов из 100-1000 клеток, которые выполняют определенные функции, «обрабатывая» определенный сигнал.
Обычно эти группы клеток напоминают вертикальные колонки. Впервые «колонки» в коре обнаружил В. Маунткасл, а в нашей стране исследованием «корковых модулей» занимался А. Б. Коган и его последователи из ростовской нейрофизиологической школы.
Особая роль этих кортикальных колонок в «ассоциативных» полях, а также во всей коре, вероятно, связана с их способностью к установлению ассоциаций, т. е. образованию временных связей, с другими колонками из других участков мозга.
11.3.2.. Миелоархитектоника
Рис. 11.4. Миелоархитектоника коры больших полушарий
Так как кора больших полушарий является высшим информационным центром нервной системы человека, то в коре имеет место интенсивный обмен нервными импульсами, как между отдельными ее участками, так и с другими отделами центральной нервной системы. Обмен информацией осуществляется по волокнам (белое вещество), соответственно, количество нервных волокон в больших полушариях очень велико. Сложно организованную систему волокон коры больших полушарий называют миелоархитектоникой. При этом всю совокупность нервных волокон можно разделить на несколько групп (рис. 11.4).
Основные пучки внутрикорковых волокон располагаются в первом слое (тангенциальные волокна), во втором слое (полоска Бехтерева), в четвертом и пятом слоях (соответственно наружная и внутренняя полоски Байарже). Эти нервные пучки отражают распространение афферентных волокон и коллате-ралей аксонов пирамидных клеток.
11. 4 Функциональное значение отдельных зон коры
Два полушария, составляющие передний мозг, действуют согласованно. Правое полушарие контролирует сенсорные и двигательные функции левой половиргы тела, а левое осуществляет аналогичный контроль над правой половиной. Связь между зрительными зонами левого и правого полушарий в норме осуществляется через мозолистое тело.
Функционально различают три разновидности коры: чувствительная
(сенсорная), двигательная (моторная) и ассоциативная (рис. 11.5).
Рис. 11.5. Разновидности коры больших полушарий по функциональному признаку
Как понятно из названия, сенсорная кора ответственна за обработку информации, поступающей от органов чувств. Именно в ней располагаются корковые отделы анализаторов человека: в затылочной области — зрительного, в височной — слухового, в теменной — кожного и т. д.
В двигательной коре располагаются первые нейроны, управляющие работой произвольных мышц человека. Каждой мышечной группе соответствует определенная область двигательной коры (рис. 11.6).
Рис. 11.6. Представительство различных мышечных групп в коре больших полушарий человека («человечек Пенфилда») [1]
Подавляющая часть корковой площади занята именно ассоциативной корой. Как полагает большинство ученых, именно там образуются ассоциативные связи между специализированными областями и интегрируется приходящая из них информация. Кроме того, здесь, как полагают, текущая информация объединяется с эмоциями и воспоминаниями, что позволяет людям думать, решать, составлять планы [1,4]. Например, считается, что ассоциативные поля теменной доли объединяют информацию, приходящую от соматосенсорной коры (сообщения от кожи, мышц, сухожилий и суставов относительно положения тела и его движений) — со зрительной и слуховой информацией, поступающей из зрительной и слуховой коры затылочной и височной долей. Как пишет Флойд Блум и его соавторы, «эта объединенная информация помогает нам иметь точное представление о собственном теле во время передвижений в окружающем пространстве. Слияние сенсорных данных с информацией, извлекаемой из кладовых нашей памяти, позволяет нам осмысленно интерпретировать специфические зрительные сигналы, звуки и тактильные ощущения. Когда что-то движущееся и пушистое коснется вашей руки, вы будете по-разному реагировать на это в зависимости от того, услышите ли вы одновременно с этим мурлыканье вашей кошки или рев медведя» [1; с. 175]. После обработки информации в ассоциативной коре интегрированная картина события передается лобной коре. Благодаря обширным двусторонним связям с лимбической системой к этой картине добавляется эмоциональный оттенок, а также информация, взятая из памяти. Другие нервные связи тоже доставляют информацию, которая позволяет лобной коре оценить текущие требования организма и окружающей среды и выбрать среди них первоочередные — решить, что лучше, а что хуже для организма в данной ситуации. Лобная кора, по-видимому, ответственна и за выбор целей, которые мы ставим перед собой на будущее, а также за нашу оценку различных обстоятельств в связи с этими целями [1].