ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 25.08.2024
Просмотров: 31
Скачиваний: 0
Для двух аллелей с полным доминированием (цвет семян гороха) возможны 3 генотипа: АА, Аа, аа и 2 фенотипа: 1-доминантный желтый (АА, Аа); 2-рецессивный зеленый (аа). Таким образом, одинаковые по фенотипу особи могут не совпадать по генотипу. Закон Харди-Вайнберга утверждает: частоты доминантного и рецессивного аллелей разных поколений идеальной популяции постоянны (идеальной можно назвать изолированную популяцию больших размеров, без новых мутаций, где спаривание происходит случайно, все генотипы одинаково плодовиты, а поколения не перекрываются). Этот закон можно выразить в уравнении Харди-Вайнберга
p2 + 2pq+q2=1, где
p2 -частота доминантных гомозигот (АА)
2pq -частота гетерозигот (Аа)
q2 -частота рецессивных гомозигот (аа)
Такое распределение возможных генотипов связано со случайным характером распределения гамет в процессе мейоза и основано на теории вероятностей, математически представляет собой квадрат уравнения вероятностей p+q=1 (уравнение вероятностей), (p+q)2=12; (p+q)(p+q)=1;
p2 + 2pq+q2=1 (уравнение Харди-Вайнберга)
Имея два уравнения для вероятностей частоты аллельных генов и наблюдая частоту рецессивных гомозигот (q2), можно вычислить число гетерозигот (2pq) – носителей скрытых генов и частоты аллельных генов (p-доминантного и q-рецессивного).
Пример решения задачи популяционной генетики.
Дано: один альбинос (аутосомно-рецессивный тип наследования) приходится на 10000 людей с нормальной пигментацией.
Найти: на сколько людей приходится 1 скрытый носитель гена альбинизма.
Решение: частота q2–рецессивных гомозигот (аа) известна из условия задачи: q2=1:10000=0,0001, необходимо найти частоту гетерозиготных носителей Х=2pq. Зная q2, можно найти частоту рецессивного аллеля q:
q
 = √q2 =
√0,0001 = 0.01
= √q2 =
√0,0001 = 0.01
Теперь, зная q, можно найти частоту доминантного аллеля p
p = 1-q = 1-0,01 = 0,99
Зная p и q, можно найти частоту гетерозиготных носителей Х=2pq
Х = 2pq=20,99 • 0,01 = 0,0198 = 0,02 или 2%.
Если 2 человека из 100 являются носителями аллельного гена альбинизма, тогда 1 носитель приходится на 50 человек.
Такие вычисления показывают чрезвычайно высокую частоту рецессивных генов в популяции при относительно редких случаях проявления наследственных заболеваний в фенотипе. Считается, что каждый человек имеет минимум 8 крайне нежелательных рецессивных генов, подавленных их нормальными аллелями.
В нацистской Германии проводились в жизнь "евгенические" программы (евгеника – наука об улучшении человеческой породы), основанные на физическом уничтожении лиц, страдающих наследственными болезнями. Несостоятельность таких способов улучшения человеческой породы ясна из закона Харди-Вайнберга. Поскольку большая часть нежелательных рецессивных генов скрыта в гетерозиготных организмах, то даже поголовное уничтожение всех выявленных рецессивных гомозигот практически не изменит числа больных в следующем поколении. Для тотального улучшения генофонда нации требуется уничтожение всех людей по обвинению в носительстве десятка дефектных генов.
Среди дефектных генов существуют летальные, которые в гомозиготном рецессивном состоянии приводят к гибели их обладателя, например, от образования внутренних спаек легких. Большая часть самопроизвольных абортов обусловлена именно летальными генами. Процент этих абортов не снижается по той же причине, что и частота наследственных болезней.
Часть дефектных генов определяет относительно стабильный процент лиц с атипичной реакцией на лекарственные средства.
На наше счастье, большая часть новых мутаций делает испорченные гены рецессивными по отношению к нормальным доминантным аллелям. Новые дефектные рецессивные гены долго (много поколений) не проявляются в популяции. Однако, в каждом следующем поколении суммируются старые и новые мутации. Возрастание такого генетического груза, пополняющегося особенно интенсивно в наше время (рост мутагенных факторов), ведет популяции к вырождению.
Генетическое вырождение людей усугубляется ослаблением естественного отбора в связи с успехами современной медицины. Альтернативой этому процессу могут быть только целенаправленные вмешательства в геном, но генная инженерия человека пока в зачаточном состоянии.
Адаптация вида (микроэволюция) – это процесс приспособления популяций к изменившимся условиям существования.
Новые сочетания генов, которые возникают преимущественно на базе комбинативной изменчивости при половом размножении, определяют уникальные фенотипы отдельных особей. Особи с неудачными (в данных условиях среды обитания) комбинациями генов и проявившимися испорченными генами, удаляются (элиминируются) из популяции, но это в соответствии с законом Харди-Вайнберга практически не снижает риск появления таких же особей в следующем поколении.
В реальных популяциях, имеющих ограниченные размеры, давление некоторых факторов среды обитания может приводить к исчезновению или значительному снижению части аллельных генов. Представьте аллельный ген, который встречается с частотой 0,01, т.е. у одного человека из 100. Если размер популяции достаточно велик (несколько миллионов особей), то случайная смерть такого человека, никак не скажется на генофонде этой популяции. Такие аллели будут тиражироваться в следующих поколениях. Если обладатель такого аллеля живет в замкнутой общине (несколько сотен), изолированной от человечества географически или социально, например, религиозный запрет на брак с иноверцами, то смерть такого человека будет означать для его популяции потерю аллеля и обеднение генофонда. Внешне это выражается в постепенном выравнивании фенотипов и росту числа больных наследственными заболеваниями.
История знает случаи вырождения династий египетских фараонов вследствие узости круга лиц, которым позволяли вступать в брак. Можно сказать, что фараоны жили в малочисленной популяции. В этом случае неизбежны близкородственные браки, ведущие к вырождению. Механизм такого вырождения сводится к увеличению коэффициента инбридинга.
Инбридинг, или близкородственное скрещивание, неизбежно сопровождается снижением жизнеспособности популяции – инбредной депрессией. При этом большая часть генов переходит в гомозиготное состояние, что манифестирует груз наследственных болезней (рис. 1).
Рисунок 1. Механизм инбредной депрессии.
отец-здоровый носитель мать-здоровый носитель
дефектных генов bce дефектных генов a g
A│ │A A│ │a
B│ │b B│ │B
C│ │c C│ │C
* * ────────────┬────────── * *
D│ │D │ D│ │D
E│ │e │ E│ │E
F│ │F ┌──────┴─────┐ F│ │F
G│ │G │ │ G│ │g
возможный генотип детей (оба здоровые
носители дефектных генов-a,b,c,e,g)
сын дочь
│A │a │A │a
│b │B │b │B
│c │C │c │C
инбридинг * * ─┐ + ┌─ * *
("кровосмешение") │D │D │ │ │D │D
│e │E │ │ │e │E
│F │F │ │A │A │ │F │F
│G │g │ │b │b │ │G │g
│ │c │c │
└ * * ┘
│D │D
│e │e
│F │F
│G │G
больной ребенок, имеющий
дефектные гены bb, cc, ee
в гомозиготном рецессивном
состоянии
Чем ближе родство лиц вступающих в брак, тем выше коэффициент инбридинга и риск рождения больных детей. Замкнутая ограниченная популяция неизбежно повышает средний коэффициент инбридинга, что ведет к ускорению темпов вырождения.
При отдаленном скрещивании – аутбридинге наблюдается прямо противоположная картина – гетерозис – или "гибридная сила". При гетерозисе заметно улучшается качество потомства. Если родители принадлежат к разным популяциям, то они имеют не совпадающий генетический груз. Другими словами, в первой популяции дрейфуют одни наследственные болезни, во второй – другие. При этом дефектные рецессивные аллели первой популяции разбавляются и маскируются нормальными доминирующими генами второй популяции.
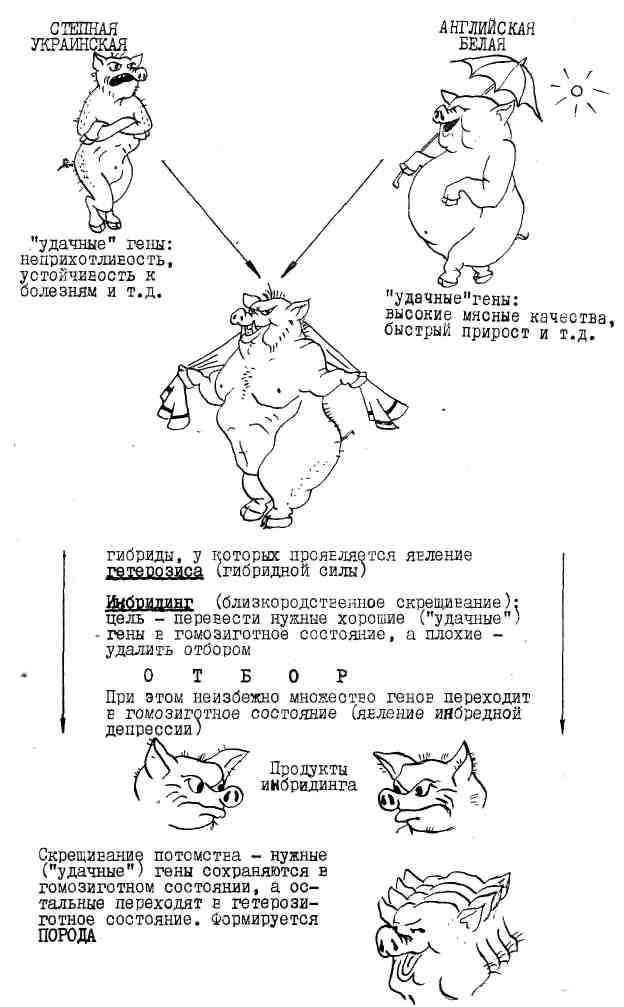
Гетерозис и последующий инбридинг широко применяют при селекционной работе с растениями и животными. На первом этапе используют отдаленную гибридизацию (гетерозис), получают высококачественное потомство, а затем путем инбридинга переводят удачные сочетания генов в гомозиготное состояние. При этом, конечно, некоторые особи рождаются с наследственными заболеваниями и их отбраковывают. Из тех особей, которым "повезло" (полезные сочетания генов перешли в гомозиготное состояние, а наследственные болезни не проявились), формируют новую породу животных или сорт растений.
Аналогичные процессы происходят в любых реальных популяциях. При этом в зависимости от влияния факторов конкретной среды обитания популяции теряют разные аллельные гены и начинают значительно отличаться друг от друга. Постепенно различия становятся препятствием к свободному скрещиванию с особями других популяций. Если скрещивание происходит, то потомство оказывается бесплодным (мулы и лошаки – гибриды лошадей и ослов, лигры - гибриды тигров и львов). На субклеточном уровне это проявляется в невозможности нормальной конъюгации гомологичных хромосом при гаметогенезе гибридов. Прекращение свободного скрещивания с появлением плодовитого потомства означает образование нового вида. Лошади и ослы действительно могли иметь общих предков, но не более примитивного, а с большим спектром аллельных генов.
Помимо гипотезы о комбинативной изменчивости с выбраковкой наиболее испорченных особей, еще одним механизмом адаптации видов к изменяющимся условиям среды обитания считается "теория эволюции". Современный неодарвинизм – это оптимистическая гипотеза. Она основана на классических представлениях об отборе особей, имеющих новые. полезные приспособления, возникшие благодаря "положительным мутациям". Положительные мутации считают молекулярной основой прогрессивных изменений и эволюции в целом. Главной проблемой этой гипотезы является противоречие с математикой из-за крайне низкой вероятности случайного возникновения новых полезных изменений генотипа. Положения неодарвинизма имеют смысл только при допущении акта разумного целенаправленного вмешательства в генетический код.
Длительное время важнейшим аргументом в пользу быстрой и целесообразной эволюции признавался факт "привыкания" микроорганизмов к лекарственным препаратам – сульфаниламидам и антибиотикам. Опыт Ледербергов показал, что это не так: на чашки Петри с питательной средой, содержащей смертельную дозу антибиотика, специальным штампом внесли отпечатки нескольких тысяч колоний бактерий, которые ранее не встречали этот антибиотик и все "должны" были погибнуть. Однако, нашли штаммы бактерий, которые были устойчивы к антибиотику и дали рост колонии микроорганизмов. При достаточно большом числе исследуемых штаммов всегда находятся устойчивые к антибиотикам. Таким образом, доказана предадаптация микробов к антибиотикам. Постепенное снижение эффективности новых противомикробных средств объясняют постепенным отбором устойчивых и вымиранием чувствительных к антибиотикам микробных штаммов.
Опыт Ледербергов показал несостоятельность доказательства гипотезы о "положительных" мутациях на основе факта появления устойчивости бактерий к действию антибиотиков.
Блок дополнительной информации.
Полиплоидия.
В современной селекции для увеличения разнообразия исходного материала все шире используется явление полиплоидии. Полиплоидией называют явление кратного увеличения набора хромосом в ядрах клеток организмов. Растения, в соматических клетках которых содержится обычный двойной набор хромосом, называются диплоидными. Если у растений набор хромосом повторяется более двух раз, они являются полиплоидными. Большинство видов пшеницы имеют 28 или 42 хромосомы и относятся к полиплоидам, хотя известны диплоидные виды с 14 хромосомами (например, однозернянка). Среди видов табака и картофеля есть виды с 24, 48 и 72 хромосомами. Полиплоидия — довольно частое явление в природе, особенно у цветковых растений (злаковых, пасленовых, сложноцветных и др.). По внешним признакам полиплоиды обычно бывают более мощными, чем диплоиды, с рослыми крепкими стеблями, крупными листьями, цветками и семенами. Это объясняется тем, что у полиплоидов клетки значительно крупнее, чем у диплоидов.
Существуют два основных типа полиплоидов: автополиплоиды и аллополиплоиды. У автополиплоидов кратно увеличен набор хромосом, характерных для данного вида (пшеница, картофель — автополиплоиды). У аллополиплоидов в геноме суммированы наборы хромосом разных видов, так как образуются аллополиплоиды при межвидовых скрещиваниях. Установлено, что явление аллополиплоидии встречается и в природе, но в основном его вызывают искусственно, удваивая число хромосом у гибридов.
Рисунок 1. Методы селекционной работы М. Ф. Иванова по выведению украинской степной белой свиньи.
Биологическая изменчивость и адаптация видов.
Эволюциионная гипотеза и креационизм - две альтернативные точки зрения на происхождение жизни и её изменения. Обе гипотезы приводят аргументы в свою пользу. Никакого отношения к вере истинно научный креационизм не имеет.
Факты не говорят сами за себя - они требуют интерпретации в рамках того или иного мировоззрения Ведущие эволюционисты - материалисты, а многие из них - ярые атеисты. Креационисты же, напротив, признают, что верят в Сотворение мира. Хотя креационисты имеют дело с теми же фактами, что и эволюционисты, они интерпретируют эти факты на основе своего мировоззрения, и приходят к обоснованным научным выводам.