ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 26.10.2023
Просмотров: 25
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Основными компонентами синтеза белка являются: аминокислоты, тРНК, аминоацил-тРНК-синтетазы, мРНК, рибосомы, источники энергии, белки - факторы инициации, элонгации и терминации и кофакторы (табл. 1).
Таблица 1
Основные компоненты белоксинтезирующей системы и их функции в процессе трансляции
| Необходимые компоненты | Функции |
| 1. Аминокислоты | Субстраты для синтеза белков |
| 2. тРНК | Выполняют функцию адапторов - приспособителей аминокислот к кодонам мРНК. Акцепторным концом (-ССА) они взаимодействуют с аминокислотами, а антикодоном - с кодоном мРНК |
| 3. Аминоацил-тРНКсинтетазы | Каждый фермент катализирует реакцию специфического связывания 1 из 20 аминокислот с соответствующей тРНК |
| 4. мРНК | Матрица содержит линейную последовательность кодонов, определяющих первичную структуру белков |
| 5. Рибосомы | Рибонуклеопротеиновые субклеточные структуры, являющиеся местом синтеза белков |
| 6. АТФ, ГТФ | Источники энергии |
| 7. Белковые факторы инициации (IF-1, IF-2, IF-3), элонгации ( EF-Tu, EF-Ts, EF-G), терминации (RF-1, Rf-2, RF-3, RRF) | Специфические внерибосомные белки, необходимые для процесса трансляции |
| 8. Ионы магния | Кофактор, стабилизирующий структуру рибосом |
| 9. Компоненты, необходимые для фолдинга и посттрансляционной модификации | Особые ферменты, кофакторы и другие компоненты для удаления начальных аминокислотных остатков, сигнальных последовательностей, протеолиза, модификации терминальных остатков, присоединения химических групп: ацетильных, метильных, карбоксильных, фосфорных, углеводных и простетических групп. |
Биосинтез белка у всех организмов происходит в 5 сновных стадий:
1.Активация аминокислот.
2.Инициация.
3.Элонгация.
4.Терминация и диссоциация рибосом.
5.Фолдинг и посттрансляционный процессинг.
6.1Активация аминокислот
тРНК выполняет адапторную роль, которая заключается: 1) в специфичном связывании с аминокислотами, 2) во включении аминокислот в белковую цепь в соответствии с матрицей мРНК.
Избирательное присоединение аминокислоты к тРНК осуществляется ферментом аминоацил-тРНК-синтетазой (рис. 7):
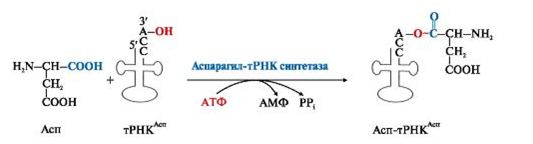
Рис. 7. Реакция активации аспартата, катализируемая аспарагил-тРНК-синтетазой
6.2Инициация трансляции
Инициация начинается с присоединения к мРНК в области «кэпа» малой субъединицы рибосомы 40S, факторов инициации (IF), инициирующей Мет-тРНКМет и ГТФ. Когда в результате движения этого комплекса по мРНК антикодон Мет-тРНКМет свяжется с инициирующим кодоном АUG, комплекс останавливается. Происходит присоединение 60S-субъединицы рибосомы, сопровождающееся гидролизом ГТФ и отделением факторов инициации. Формируется 80S-рибосома с двумя активными центрами: Р (пептидильным) центром, в котором находится Мет-тРНКМет, и А(аминоацильным) центром, в область которого поступает первый смысловой кодон мРНК (рис. 8.).
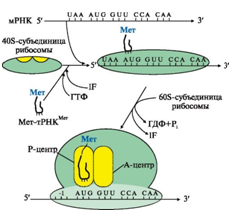
Рис. 8. Инициация белкового синтеза
6.3. Элонгация трансляции
Стадия элонгации включает три последовательных этапа: 1) связывание аа-тРНКаа в А-центре, 2) образование пептидной связи, 3) транслокация (рис. 9).
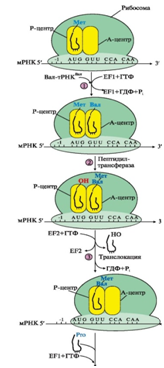
Рис. 9. Элонгация полипептидной цепи:
1 - связывание аа-тРНКаа
в А-центре требует затраты энергии ГТФ и участия фактора элонгации EF1 (на схеме аа-тРНКаа - Вал-тРНКВал); 2 - образование пептидной связи катализирует пептидилтрансфераза, активный центр которой формируется рРНК, входящей в состав большой субъединицы рибосомы; 3 - перемещение рибосомы по мРНК на один кодон в направлении от 5'- к 3'-концу идет с использованием энергии ГТФ (транслокация) и при участии фактора EF2.
6.4 Терминация трансляции
Терминация трансляции происходит после включения в А-центр одного из стоп кодонов: UAG, UGA, UAA (рис. 10).
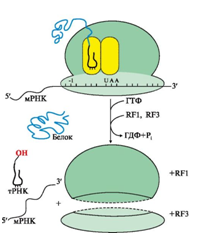
Рис.10. Терминация синтеза белка
7. Общая теория регуляции синтеза белка по Ф. Жакобу и Ж. Моно. Ингибиторы синтеза белка. Механизм действия дифтерийного токсина на биосинтез белка
Механизм регуляции генетического кода был открыт французскими учеными Ф. Жакобом и Ж. Моно в 1961 г. на бактериях. E. coli и получил название механизма индукции-репрессии. В основе теории лежат следующие понятия:
-оперон – группа тесно связанных между собой генов, которые программируют образование структурных белков и ферментов в клетке,
- конституитивные гены – те, которые активны в клетках всегда, независимо от ее активности,
-индуцибельные гены – те, которые считываются при необходимости,
-ген-регулятор – ген, регулирующий работу оперона, но не входящий в его состав. Он синтезирует белок-регулятор (чаще называемый белок-репрессор), который может быть в активной или неактивной форме,
-ген-оператор – участок ДНК, способный связываться с белком-регулятором, и "решающий" нужно работать РНК-полимеразе или нет.
Если оперон регулируется по механизму индукции (например, лактозный оперон), то в отсутствии индуктора (лактозы) белок-репрессор связан с оператором. Комплекс белок-репрессор-оператор препятствует связыванию РНК-полимеразы с промотором, и транскрипция структурных генов оперона не идет. Когда концентрация индуктора в клетке возрастет, то он присоединится к белку-репрессору, который имеет центр для связывания индуктора, изменяет его конформацию и снижает сродство к оператору. Комплекс индуктор-белок-репрессор теряет сродство к ДНК и уходит в цитозоль клетки. РНК-полимераза связывается с промотором и транскрибирует структурные гены. Идет синтез белков, закодированных в данном опероне.
Лекарственная регуляция
Ингибирование
1. Гетероциклические соединения доксорубицин, дауномицин и актиномицин D обладают способностью встраиваться между нитей ДНК между двумя соседними парами оснований Г-Ц, препятствуя движению РНК-полимеразы.
2. Рифампицин связывается с β-субъединицей РНК-полимеразы прокариот и ингибирует ее. Благодаря такой избирательности действия рифампицин действует только на бактерии и является препаратом для лечения туберкулеза.
3. α-Аманитин, октапептид бледной поганки (Amanita phalloides) блокирует РНК-полимеразу II эукариот и предотвращает продукцию мРНК.
4. Энтеротоксин возбудителя дифтерии Сorynebacterium diphteriae ингибирует синтез белков в клетках слизистой оболочки зева и гортани. В цитоплазме клеток под влиянием протеолитических ферментов токсин расщепляется на два фрагмента, один из которых является АДФ-рибозилтрансферазой. Этот фермент катализирует реакцию:
FE-2 + NAD → АДФ-рибозил-FE-2 + никотинамид + Н+
Субстратом реакции является фактор элонгации FE-2, модификация которого нарушает транслокацию рибосом, прекращает биосинтез белков в инфицированных клетках и вызывает их гибель.
Активация
Активация транскрипции используется в клинике намного реже и заключается в применении аналогов стероидных гормонов для достижения анаболического эффекта в органе-мишени.