Файл: Аминоацилирование трнк механизм действия аминоацилтрнксинтетаз.docx
Добавлен: 26.10.2023
Просмотров: 26
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Санкт-Петербургский Государственный Химико-Фармацевтический Университет
Кафедра биологической химии
Реферат
На тему: «Аминоацилирование тРНК: механизм действия аминоацил-тРНКсинтетаз»
Подготовила:
Студентка 1 курса
фармацевтического факультета
Группа ФС-3322
Ушакова Анастасия Павловна
Дата:____________Подпись___________
Научный руководитель:
Доцент Ольга Михайловна Спасенкова
Дата:____________Подпись___________
Санкт-Петербург
2023
Содержание
1. Роль тРНК в трансляции .......................................................................стр.3
2. Аминоацил тРнк синтетазы....................................................................стр.3
3. Белоксинтезирующая система клетки ...................................................стр.5
4. Инициация трансляции............................................................................стр.6
5. Элонгация трансляции .............................................................................стр.9
6. Терминация трансляции........................................................................стр.11
7. Эффективность трансляции.................................................................стр. 12
8. Посттрансляционные модификации полипептидной цепи................стр 13
Литература
1. Роль тРНК в трансляции
В белоксинтезирующей системе тРНК выполняет следующие три важные функции:
а) акцепторную (с помощью специфического фермента ами- ноацил-тРНК-синтетазы присоединяет на одном из концов своей молекулы соответствующую аминокислоту, в результате чего возникает комплекс ами- ноацил-тРНК);
б) транспортную (доставляет аминокислоту в форме аа-тРНК в рибосому для включения ее в растущую полипептидную цепь);
в) адапторную (с помощью своего антикодона специфически взаимодействует с комплементарным ему кодоном мРНК и таким образом обеспечивает необходимую последовательность включения аминокислот в синтезируемую полипептидную цепь в соответствии с программой, заданной мРНК).
Благодаря адапторной функции, тРНК «дешифрует» генетический код в РНК-матрице и переводит его в аминокислотный код белка. Реализация всех этих функций возможна благодаря уникальной структуре молекул тРНК.
2. Аминоацил тРнк синтетазы
Это особый класс ферментов, катализирующих совокупность реакций, составляющих первую стадию биосинтеза белка – строго специфическое соединение аминокислоты с соответствующей ей тРНК. Для каждой АК существует специфическая аа-тРНК-синтетаза. Клетка содержит, по крайней мере, 20 типов аа-тРНК-синтетаз, обладающих специфичностью не только в отношении аминокислоты, но и тРНК. Из-за единообразия функций все тРНК имеют очень сходную пространственную структуру. Поэтому распознавание аа-тРНК-синтетазой своей тРНК должно базироваться на очень тонких различиях строения отдельных тРНК.
Аминоацил-тРНК-синтетазы имеют два активных центра: один небольших размеров, связывающий аминокислоту; и второй, протяженный, для точного выбора тРНК. АТР связывается в активном центре фермента через ион Mg2+ с имидазольным радикалом His.
Ферменты, выделенные из одной и той же клетки, но специфичные к разным АК, существенно отличаются по субъединичной структуре и молекулярной массе. Молекулярная масса большинства аа-тРНК-синтетаз составляет примерно 100 кДа, иногда 200 кДа и больше. Эти ферменты являются димерами или тетрамерами, очень редко имеют одну полипептидную цепь, содержащую до 1000 аминокислотных остатков (изолейцил-тРНК-синтетаза, аланил-тРНК-синтетаза). Структура аа-тРНК-синтетаз обеспечивает точную установку тРНК относительно фермента, в результате чего участок аа-тРНК- синтетазы, несущий активированную АК, располагается рядом с 3′-концевым аденозином.
Аминоацил-тРНК-синтетазы работают очень точно: ошибочное аминоацилирование in vivo встречается только приблизительно в одном случае из 10000 циклов этой реакции. Именно в виде аа-тРНК аминокислота непосредственно вовлекается в биосинтез белка, осуществляемый белоксинтезирующей системой клетки. Аминоацил-тРНК-синтетазы очень медленные ферменты, число их оборотов составляет 50-500 каталитических актов в минуту.
Аминоацил-тРНК-синтетазы делят на два класса: класс I (ферменты, переносящие остаток аминокислоты на 2′-ОН группу рибозы); класс II (ферменты, переносящие остаток аминокислоты на 3′-ОН группу концевой рибозы тРНК).
Аминоацил-тРНК-синтетазы существуют в виде высокомолекулярных комплексов – кодосом. Их молекулярная масса 1,4 мДа. Они включают несколько аа-тРНК-синтетаз и ферменты, модифицирующие аа-тРНК- синтетазы и регулирующие их активность. Это протеинкиназы, метилтрансферазы, фосфопротеинфосфатазы и др.
Аминоацил-тРНК-синтетазам свойственны и неканонические функции. В частности, некоторые митохондриальные аа-тРНК-синтетазы проявляют сплайсирующую активность и участвуют в процессинге мРНК. Механизм этого явления неясен.
Как осуществляется узнавание тРНК аа-тРНК-синтетазами? Элементы узнавания тРНК, по-видимому, отличаются у про- и эукариот. Это было показано в экспериментах: многие бактериальные тРНК плохо аминоацилируются аа-тРНК-синтетазами млекопитающих, и наоборот, тРНК млекопитающих служат плохими субстратами для аа-тРНК-синтетаз
Е.соli.
Элементами узнавания у прокариот и дрожжей служат:
а) антикодон. (главный элемент узнавания для большинства тРНК);
б)Нуклеотиды акцепторного стебля; в) вариабельная шпилька (в том случае, если она достаточно длинная).
Нуклеотиды антикодона остаются основными элементами узнавания как в про-, так и в эукариотических системах. В других случаях элементами узнавания служат несколько оснований в разных участках молекулы тРНК.
Активный центр фермента высокоспецифичен в отношении субстрата, но пределы точности все же существуют. Фермент достаточно легко отличает аминокислоты с сильно различающимися свойствами, но ему трудно отличить похожие аминокислоты, например валин и изолейцин.
В случае ошибочного аденилирования изолейцил-специфический фермент гидролизует валил-АМР, в то время как изолейцил-АМР (возможно изза гораздо больших размеров) в этот центр не входит. Благодаря этому уменьшается ошибка присоединения изолейцина до 1 на 60000. Не все аа- тРНК-синтетазы имеют такой механизм коррекции. Он нужен только для распознавания похожих аминокислот.
3. Белоксинтезирующая система клетки
В состав белоксинтезирующей системы входят следующие компоненты:
1) рибосомные субъединицы 30S и 50S, образующие у прокариот рибосому 70S, или субъединицы 40S и 60S, образующие у эукариот рибосому
80S;
2)мРНК;
3)полный комплект аминоацил-тРНК, для образования которых необходимы аминокислоты, аминоацил-тРНК-синтетазы, тРНК и АТР;
4)инициаторная аа-тРНК. У прокариот – формилметионил-тРНК, у эукариот – метеонил-тРНК;
5)белковые факторы инициации трансляции. У прокариот – IF-1, IF-2, IF-3, у эукариот 9 факторов: еIF-1, еIF-2, eIF-3, eIF-4A, eIF-4B, eIF-4C, eIF4D, eIF-5, eIF-6. Для инициации трансляции у эукариот абсолютно необходимы еIF-2, eIF-3 и eIF-5, остальные факторы усиливают функции этих трех;
6)белковые факторы элонгации: у прокариот – EF-Tu, EF-Ts, EF-G (Tu, Ts, G), у эукариот – EF-1 (аналог Tu), EF-2 (аналог EF-G);
7)белковые факторы терминации (или освобождения): у прокариот –
RF-1 (R1), RF-2 (R2), RF-3 (S), у эукариот – еRF (для проявления активности ему необходим GTP);
8)некоторые другие факторы, еще недостаточно хорошо изученные (факторы диссоциации, ассоциации, высвобождения и др. белки);
9)GTP;
10)неорганические катионы Mg2+ или Са2+ и одновалентные ( К+ или NH4+) в определенной концентрации.
В ходе синтеза белка информация, закодированная в мРНК, «читается»
внаправлении от 5 - к 3 -концу, обеспечивая синтез полипептида от N- к С-концу. У прокариот мРНК полицистронна, синтез белка на данной матрице начинается еще до того, как заканчивается процесс транскрипции (транскрипция и трансляция в прокариотических клетках сопряжены в пространстве и во времени). В отличие от прокариот, у эукариот мРНК моноцистронны и каждая мРНК кодирует строение только одной полипептидной цепи. В клетках эукариот синтез белка и транскриция разобщены. Транскрипция осуществляется в ядре клетки, трансляция в цитоплазме, куда из ядра поступают «зрелые», функционально активные молекулы мРНК.
4. Инициация трансляции
Рибосома должна узнать первый триплет кодирующей последовательности и там начать трансляцию. Необходима абсолютно точная инициация, поскольку правильность трансляции мРНК рибосомой зависит от правильной рамки считывания. Если произойдет сдвиг рамки считывания, аминокислотная последовательность полипептида, синтезированного в этом случае, окажется ошибочной, а образовавшийся продукт будет не способен выполнять функции белка, закодированного в данном гене.
В прокариотических клетках инициация трансляции является внутренней. Это означает, что рибосомная 30S-частица присоединяется к участку мРНК, содержащему инициирующий кодон AUG. Не имеет значения, на каком расстоянии от 5 -конца мРНК кодон находится. Для прокариот этот способ инициации трансляции является оптимальным, поскольку он обеспечивает инициацию трансляции сразу нескольких цистронов внутри полицистронных мРНК.
В расположении 30S-субчастицы на мРНК важную роль играет последовательность Шайна-Дальгарно (рис. 31.1), являющаяся элементом 5 - нетранслируемой области прокариотической мРНК. Инициаторной аминоа- цил-тРНК у прокариот является fMet-тРНКfMet, взаимодействующая антикодоном UAC с колоном AUG на мРНК по принципу комплементарности.

Рис. 31.1. Присоединение 30S-частицы и fMet-тРНКfMet к МРНК у прокариот
У эукариот инициация трансляции называется терминальной инициацией. В этом случае 40S-частица сначала присоединяется к 5 -концу мРНК, затем движется по мРНК до тех пор, пока не встретит инициирующий кодон. Этот процесс называют сканированием мРНК; он требует затраты энергии и является АТР-зависимым. В АТР-зависимом расплетании вторичной структуры мРНК и сканировании ее первичной структуры участвует специальный эукариотический фактор инициации eIF4, обладающий АТР-азной и хеликазной активностью. Когда рибосомная частица 40S встречается с инициирующим кодоном, антикодон инициаторной Met-тРНКMet взаимодействует с ним и сканирование прекращается (рибосомная частица нашла начало кодирующей последовательности мРНК).
Старт-кодоном в мРНК эукариот также является кодон AUG, но данный триплет может находиться в любой части мРНК, т.к. кодирует аминокислоту – метионин. В связи с этим существуют две различные тРНК, специфичные для метионина. Обе обладают одним и тем же кодоном, но одна используется только для инициации трансляции, а другая – только для включения метионина в процессе элонгации. Инициаторная тРНК имеет структурные особенности, которые распознаются инициаторным белком, или фактором инициации еIF-2, осуществляющим ее доставку к формирующемуся инициаторному комплексу. Мet-тРНК, участвующие в элонгации, опознаются другим цитоплазматическим фактором, который и доставляет ее к рибосоме. Этот фактор не связывается с инициаторной Met-тРНК.
В цитоплазме существует фонд свободных 30S- и 50S-субчастиц рибосомы. Белковый фактор IF-3 связывается с 30S-субчастицей и предупреждает реассоциацию рибосомных частиц при инициации трансляции. Этот фактор должен быть высвобожден до того, как 50S-субчастица сможет присоединиться.
Белковые факторы IF-1 и IF-2, связываясь с 30S-субчастицей, участвуют в процессе инициации и освобождаются в цитоплазму, чтобы функциони-
ровать вновь в новом акте инициации. Роль IF-1 точно неизвестна, а IF-2 необходим для связывания инициаторной fmet-тРНКfmet (рис. 31.2).
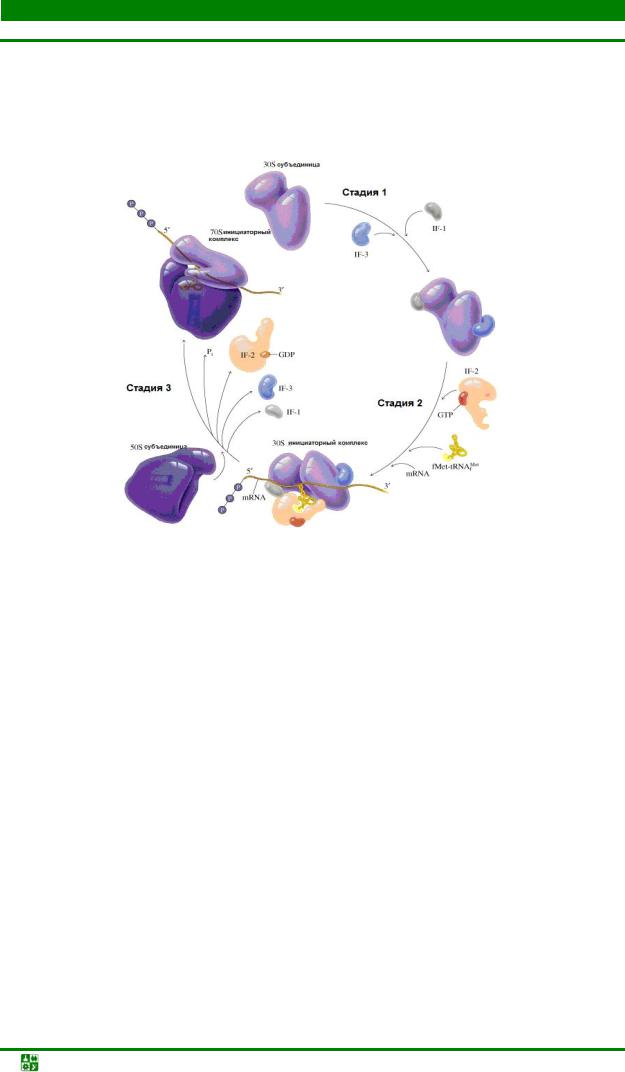
Рис.31.2.Образование инициирующего 70Sкомплекса у прокариот
Участки Р и А окончательно формируются только при присоединении 50S-субчастицы. Во время синтеза белка Р-сайт оккупируется молекулой тРНК, на которой находится растущая полипептидная цепь; А-сайт занят аминоацил-тРНК. Растущая полипептидная цепь проходит через туннель на большой субчастице (рис. 31.3).
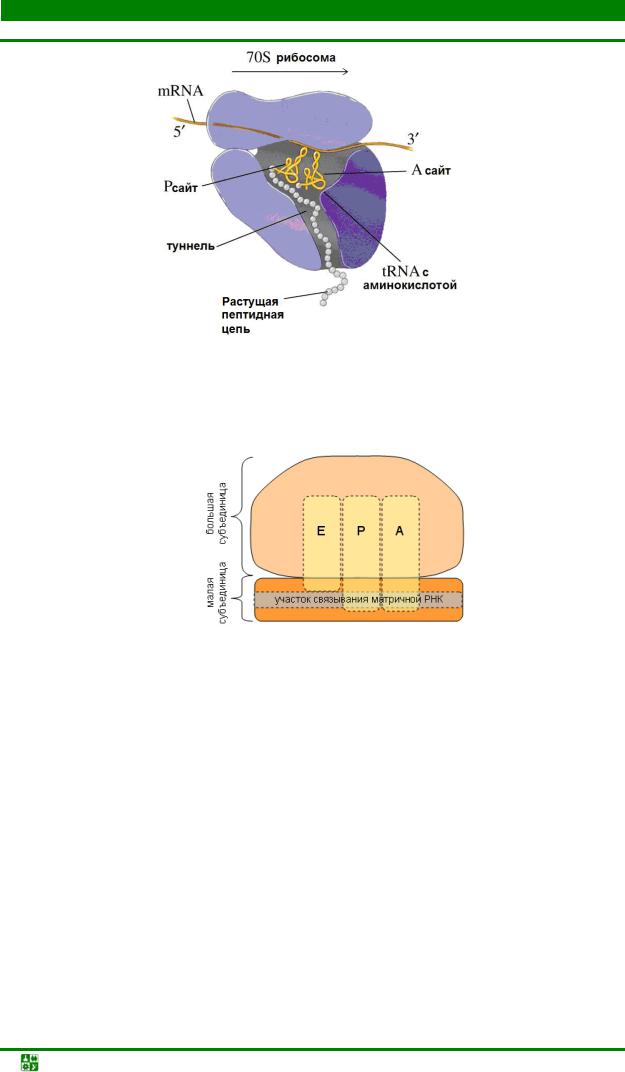
Рис.31.3. 70Sинициирующий комплекс у прокариот
Кроме А-сайта (акцепторного) и Р-сайта (донорного) в 70S-комплексе формируется Е-сайт, с которого уходит деацилированная (без аминокислоты)
тРНК (рис. 31.4).
Рис. 31.4. Схема РНК-связывающих участков рибосомы: А аминоацил-тРНК- связывающий участок; Р пептидил-тРНК-связывающий участок, Е участок выхода тРНК (от англ. exit)