Файл: Аминоацилирование трнк механизм действия аминоацилтрнксинтетаз.docx
Добавлен: 26.10.2023
Просмотров: 27
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
5. Элонгация трансляции
Выделяются следующие три стадии элонгации трансляции:
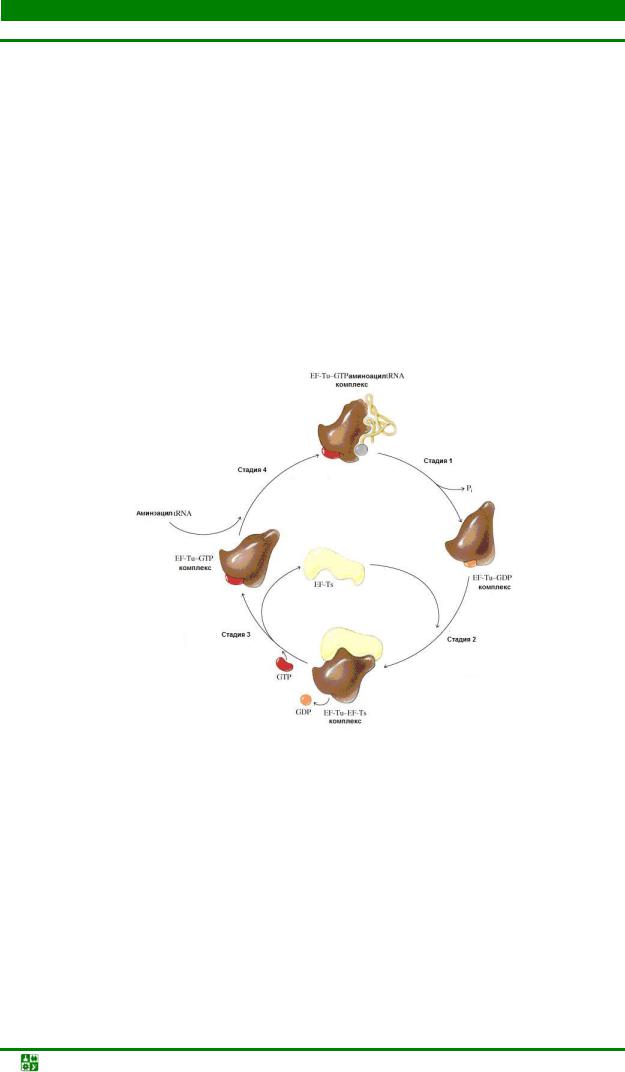
Стадия 1. Кодонспецифический выбор аа-тРНК. Требует обязательного участия белковых факторов Tu, Ts и GTP. Реакции протекают в следующей последовательности. Вначале в результате взаимодействия между белковым фактором EF-Tu и GTP образуется относительно нестабильный комплекс EF- Tu-GTP. Образованный комплекс неспецифически связывает одну молекулу любой аа-тРНК:
EF-Tu-GTP + аа-тРНК → аа-тРНК-EF-Tu-GTP
Далее происходит связывание аа-тРНК-EF-Tu-GTP на R-участке, которое происходит, вероятно, еще во время предыдущего рабочего цикла рибосомы. Комплекс аа-тРНК- EF-Tu-GTP некоторое время удерживается на участке предварительного узнавания (до переноса на А-сайт). Механизм этого переноса остается неизвестным.
70S-мРНК + аа-тРНК- EF-Tu-GTP → 70S-мРНК-аа-тРНК- EF-Tu-GTP
Следующий этап гидролиз GTP до GDP, который остается в комплексе с EF-Tu и Н3РО4. Они высвобождаются из рибосомы. В цитоплазме происходит при участии фактора EF-Ts возвращение EF-Tu-GDP в исходное состояние:
EF-Tu-GDP + EF-Ts + GTP → EF-Tu-GTP + EF-Ts + GDРНа рис. 31.5 приведен циклический процесс регенерации EF-Tu-GDP.
Стадия 2. Транспептидация. Свободная NH2-группа аа-тРНК ориентируется рядом с этерифицированным карбоксилом пептидил-тРНК. Такое пространственное сближение субстратов пептидилтрансферазного центра (ПТЦ) является необходимым и достаточным условием образования между ними пептидной связи. Эта реакция катализируется самим ПТЦ. В процессе транспептидации пептидил, связанный через СОО-группу с тРНК в донорном центре, покидает свою тРНК (она становится деацилированной) за счет замыкания пептидной связи переносится на NH2-группу аминокислоты аатРНК.
В результате единичной транспептидации пептидил удлиняется на одни аминокислотный остаток. Необходимая для этогог энергия запасена в сложноэфирной связи пептидила (fmet) и концевого аденозина тРНК
(рис. 31.6).
Важным следствием транспептидации является резкое снижение прочности удержания измененных субстратов ПТЦдеацилированную тРНК в донорном участке и пептидил-тРНК в акцепторном участке, что необходимо для прохождения следующей стадии цикла – транслокации.
Стадия 3. Транслокация. После замыкания петидной связи донорная тРНК, лишившаяся пептидила, занимает донорный участок, а пептидил оказывается связанным с акцепторной тРНК в А-участке. Такое состояние рибосомы и пептидил-тРНК называется претранслоцированным.
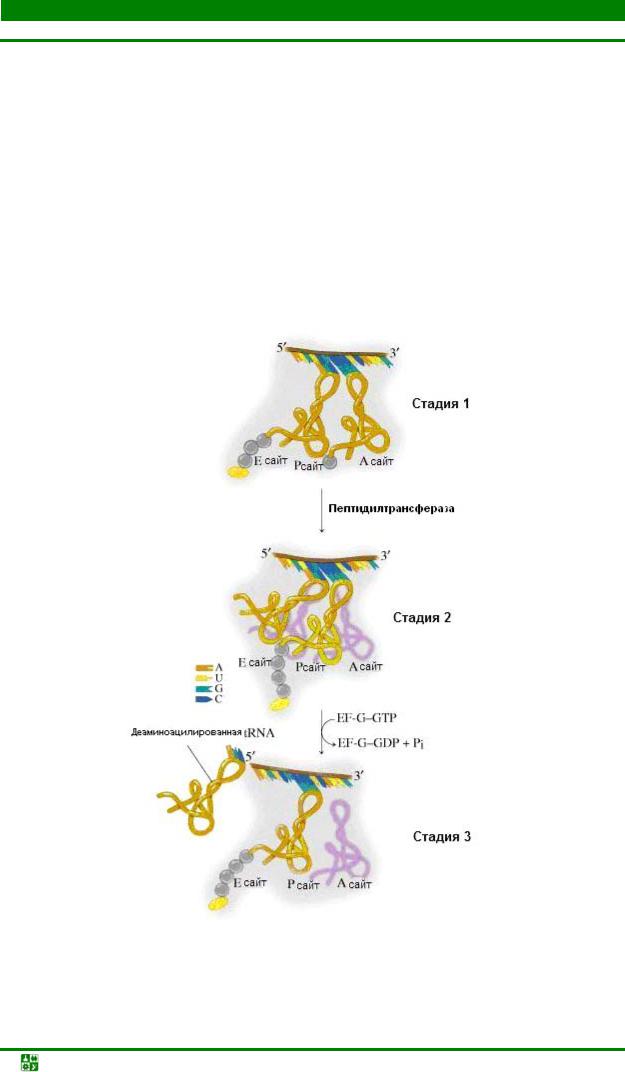
Рис.31.6. Элонгация трансляции у прокариот
Чтобы рибосома могла присоединить очередную аа-тРНК и образовать следующую пептидную сявзь, в ней должны произойти пространственные перемещения некоторых компонентов. Этот процесс получил название транслокации. Она включает следующие события:
1). Перемещение пептидил-тРНК с акцепторного на донорный участок
– транслокация тРНК;
2). Вытеснение деацилированной тРНК из Р-участка;
3). Перемещение рибосомы вдоль мРНК в направлении 5′ → 3′ на один кодон и установка в акцепторном участке нового кодона – транслокация рибосомы или транслокация мРНК.
Во время транслокации очередная аа-тРНК, вероятно, перемещается из R-участка в А-участок (рис. 31.6).
Транслокация в клетке происходит с участием белкового фактора EF-G связанного с GTP (транслоказа). Для удаления EF-G из рибосомы, которое происходит сразу после транслокации, необходим нидролиз GTP. Такое состояние рибосомы называется посттранслоцированным. Рибосома способна повторить весь цикл снова.
Таким образом, на этап элонгации затрачивается две молекулы GTP.
6. Терминация трансляции
Терминация трансляции – это процесс завершение синтеза п/п цепи и освобождение ее из связи с последней тРНК и рибосомой. Сигналом о завершении трансляции является один из трех бессмысленных кодонов: UAA, UAG, UGA. Помимо терминирующих кодонов в терминации трансляции участвуют три белковых фактора – RF-1, RF-2, RF-3.
Основные стадии терминации:
1). Узнавание терминирующего кодона.
Рибосома должна находиться в посттранслоцированном состоянии. ТТ начинается с того, что в А-сайт поступает один из терминирующих кодонов. Поскольку этим кодонам не соответствует какая-либо аа-тРНК, с этим участком связывается один из факторов терминации – RF-1 или RF-2. Эта реакция стимулируется фактором RF-3.
2). Гидролиз сложноэфирной связи между С-концом пептидила и ССАконцом донорной тРНК.
Эта реакция осуществляется ПТЦ рибосомы. Факторы терминации делают его способным переносить пептидил на Н2О, вследствие чего пептидил отделяется от рибосомы, но мРНК и деацилированная тРНК продолжают на ней удерживаться. На этом процесс терминации трансляции заканчивается, и все последующие стадии необходимы только для подготовки рибосомы к следующей трансляции.
3). Освобождение рибосомы из комплекса с мРНК и тРНК. Фактор ERF, фактор элонгации EF-G и GTP. Механизм неизвестен. 4). Диссоциация рибосомы 70S Эта стадия протекает с участием IF-3, который специфически взаимодействуя с 30S, способствует ее отделению от 50S.
7. Эффективность трансляции
Под эффективностью трансляции понимают скорость включения аминокислот в полипептидной цепи. В оптимальных условиях время, необходимое для синтеза п/п цепи, включающей 300-400 а.о., составляет у Е.соli 10-20 сек, 30-40 (15-20) а.о./сек. У эукариот за секунду включается в п/п цепь 10 а.о.
Следовательно, элонгация п/п цепи небольших размеров продолжается менее 10-30 секунд. Скорость синтеза белка в ретикулоцитах составляет около 1 триплета в секунду и около 7-10 триплетов в секунду у Е.соli.
В среднем на каждые 104 аминокислот включается одна неправильная АК и, следовательно, одна ошибка приходится на каждые 25 синтезированных белков, имеющих средний размер (примерно 400 АК).
Точность процесса трансляции зависит от надежности двух механиз-
мов.
1). Связывания каждой аминокислоты с соответствующей молекулой тРНК.
2) Спаривание кодонов мРНК с антикодоном тРНК. Оба механизма нуждаются в затрате энергии.
У аа-тРНК-синтетазы есть 2 центра, один из них отвечает за присоединение аминокислоты к тРНК, второй – гидролитический, отвечающий за удаление неправильной АК, присоединившейся к тРНК. Этот центр распознает неправильно включенную аминокислоту.
На включение одной аминокислоты в растущую п/п цепь затрачивается 4 макроэргические связи: 2 из АТР в ходе активации аминокислоты (реакция, катализируемая аа-тРНК-синтетазой) и 2 молекулы GTP: одна на связывание аа-тРНК в А-сайте рибосомы, вторая - на транслокацию. Кроме того, необходимо учитывать использование еще двух молекул GTP у прокариот, затрачиваемых на стадию инициации и терминации трансляции. У эукариот на стадию инициации (сканирование мРНК) затрачивается еще и молекула АТР.
8. Посттрансляционные модификации полипептидной цепи
Пептидная цепь, растущая в процессе трансляции, принимает вторичную и третичную структуру в результате сложного многоступенчатого процесса, идущего во времени. Для образования правильной структуры с еще не свернувшейся пептидной цепью связываются специальные белки – шапероны. Шапероны обладают сродством к экспонированным гидрофобным участкам п/п цепи. Связывание с шаперонами препятствует агрегации с другими белками и тем самым создает условия для нормального сворачивания растущего пептида. Взаимодействие с шаперонами – процесс энергозависимый: при освобождении шаперонов гидролизуется АТР (
рис. 31.7).
Шапероны принадлежат к трем белковым семействам, т.н. белкам теплового шока – heat shok proteins (hsp60, hsp70, Hsp90). Свое название эти белки получили потому, что их синтез возрастает при повышении температуры и других формах стресса. При этом они выполняют функцию защиты белков клетки от денатурации. Белки – представители семейства hsp70 – связываются на начальной фазе образования растущего пептида. Одни из них контролируют процесс сворачивания белка hsp60 охватывают синтезированный полипептид наподобие бочонка, тем самым обеспечивая условия для принятия правильной конформации.
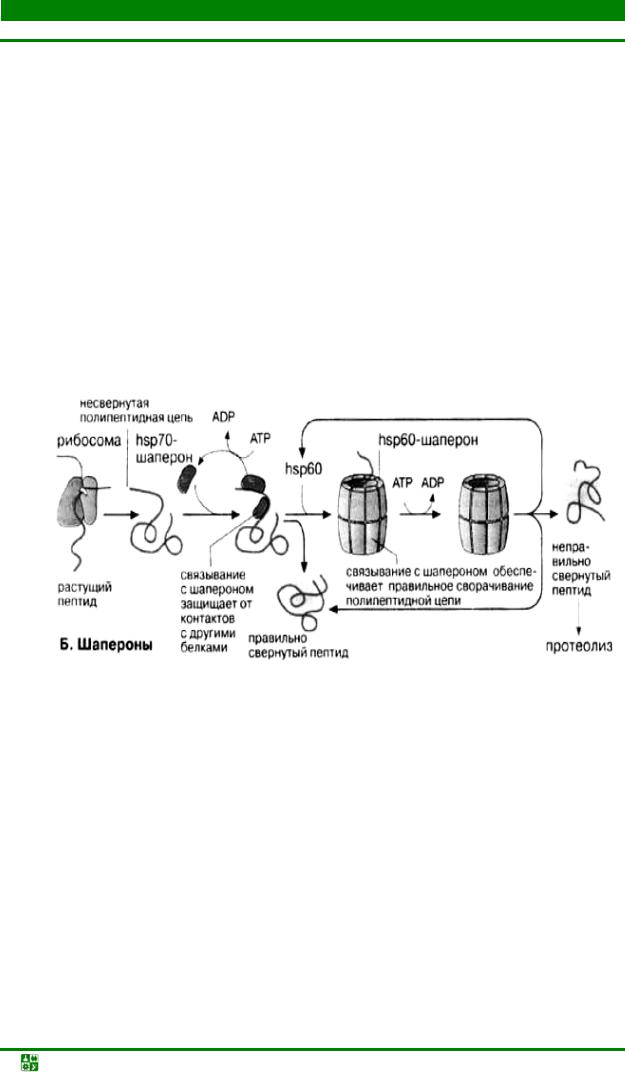 Рис. 31.7. Роль шаперонов в фолдинге полипептидной цепи
Рис. 31.7. Роль шаперонов в фолдинге полипептидной цепиПревращение линейной немодифицированной пептидной цепи в полноценный функциональный белок (созревание) осуществляется в результате многостадийного процесса, который начинается сразу же после начала трансляции и протекает в просвете ЭР (рис. 31.8).
Прежде всего соответствующая пептидаза отщепляет сигнальный пептид (1). Фермент узнает точку расщепления в составе специфической N- концевой последовательности белка. Путем окисления боковых цепей цистеина образуются дисульфидные мостики, правильность положения которых контролируется протеиндисульфид-изомеразой (2). Пептидилпролилизомераза контролирует цис-транс-изомеризацию Х-Рrо-связей в синтезируемом пептиде (3). Трансгликозидазы переносят олигосахариды в блоке с долихолом (длинноцепочечным изопреноидом) на определенные остатки аспарагиновой кислоты в белке, тем самым осуществляя N-гликозилирование белка (4). Гликозидазы «подстригают» олигосахариды, отщепляя избыточные остатки глюкозы и маннозы (5).
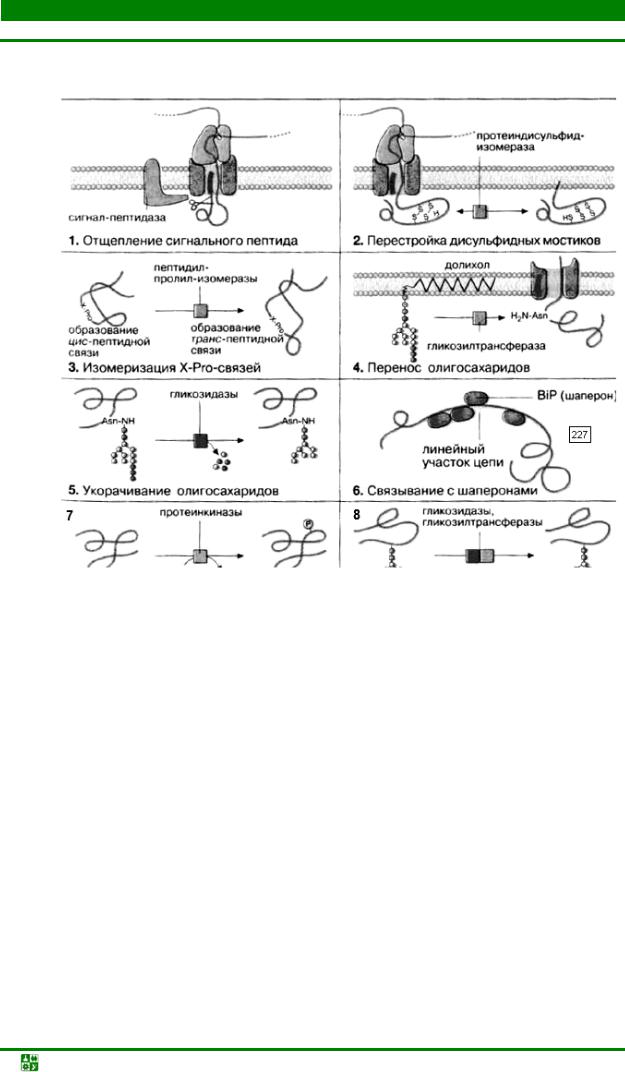 Рис.31.8. Примеры посттрансляционной модификации полипептидной цепи
Рис.31.8. Примеры посттрансляционной модификации полипептидной цепиДля того чтобы растущая полипептидная цепь могла свернуться необходимым образом, с еще линейным участком цепи временно связываются шапероны (6). Эти белки направляют процесс свертывания цепи путем подавления нежелательных побочных взаимодействий. Наиболее важным шапероном, присутствующим в просвете ШЭР. является
белок связывания (BiP, от англ. binding protein). Когда вновь образованный белок приобретает правильную вторичную и третичную структуру и остатки глюкозы удалены полностью, он с помощью транспортных везикул перемещается в аппарат Гольджи. В аппарате Гольджи осуществляются следующие ферментативные стадии модификации белка: фосфорилирование (7) и отщепление с последующим переносом (перегруппировка) остатков сахаров с помощью гликозидаз и гликозилтрансфераз (8). Эта модификация имеет целью образование специфической олигосахаридной структуры в гликопротеинах.
Список используемой литературы
1.Альбертс Б., Брей Д., Льюис Дж. И др. Молекулярная биология клетки:В 3-х т. – М.: Мир, 1994. Т.1. - с. Т.2. - с. Т.3. - с.
2.Березов Т.Т., Коровкин Б.Ф. Биологическая химия: Учебник. – 3-е изд.. перераб. и доп. – М.: Медицина, 1998. – 704 с.: ил. – (Учеб. лит. для студентов мед. вузов).
3.Биохимия: Учебник /Под ред. Е.С. Северина. – М.: ГЭОТАР-МЕД, 2003. – 784 с.: ил. (Серия «XXI век»).
4.Биохимия. Краткий курс с упражнениями и задачами /Под ред Е.С. Северина, А.Я Николаева. – М.: ГЭОТАР-МЕД, 2001. – 448 с.: ил. (Се-
рия «XXI век»).
5.Граник В.Г. Метаболизм эндогенных соединений: Монография. – М.: Вузовская книга, 2006. – 528 с.: ил.
6.Кнорре Д.Г., Мызина С.Д. Биологическая химия: Учеб. для хим., биол. и мед. спец. вузов. – 2-е изд., перераб. и доп. – М.: Высш. шк., 1998. – 479 с.: ил.
7.Кольман Я., Рэм К-Г. Наглядная биохимия. – М.: Мир, 2000. – 469 с.
8.Коничев А.С., Севастьянова Г.А. Молекулярная биологи: Учеб. для студ. пед. вузов. – М.: Издательский центр «Академия», 2003. – 400 с.
9.Молекулярная биология: Структура и биосинтех нуклеиновых кислот: Учеб. для биол. спец. вузов/В.А. Агол, А.А. Богданов, В.А. Гвоздев и др.; Под ред А.С. Спирина. – М.: Высш.шк., 1990. – 352 с.