ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 16.11.2021
Просмотров: 169
Скачиваний: 1
Тема: Регулятори росту і розвитку рослин (РРР).
1. Фітогормони в онтогенезі рослин;
2. Використання регуляторів росту рослин в практиці рослинництва;
3. Класифікація фітогормонів та методи їх отримання;
4. Основні типи РРР, їх фізіологічна дія та використання.
1.Регулятори росту рослин (РРР) – це природні або синтетичні органічні сполуки, які активно впливають на обмін речовинвищих рослин, регулюють фізіологічні та морфогенетичні програми росту і розвитку рослинного організму. Регулятори росту, які продукуються самою рослиною, називаються фітогормонами. За типом дії РРР поділяють на стимулятори та інгібітори. Проте такий поділ умовний, оскільки більшість із них в низьких концентраціях стимулює, а у високих - пригнічує ті чи інші процеси (2,4Д у високих концентраціях є сильний гербіцид, а в низьких -синтетичний регулятор росту рослин).
Фітогормони переміщуються по рослині і впливають на ріст і диференціацію тих тканин і органів, куди потрапляють. Таким чином, фітогормони – солуки, за допомогою яких здійснюється взаємодія клітин, тканин і органів рослин. Вони синтезуються і функціонують в мікрокількостях і на відміну від інших метаболітів, у тому числі вітамінів, здатні викликати в рослині формоутворюючі процеси (ріст коренів, пагонів, утворення квіток, плодів, тощо).
2. З експертних оцінок наукових закладів відомо, що продуктивність нових сортів і гібридів с.г. культур нижча їх генетичної можливості на 40 і більше відсотків. У зв’язку з цим господарства, як правило, висівають насіння польових культур з високими нормами висіву, що призводить до перевитрат високоякісного зерна. Цьому можна на сьогодні запобігти , якщо використовувати синтетичні регулятори росту, які допомагають рослинам протистояти несприятливим факторам і проявити сві генетичний потенціал.
У практиці рослинництва широко використовують синтетичні РРР для підвищення енергії проростання і польової схожості насіння, запобігання виляганню хлібів, регуляції плодоношення, підвищення врожайності, приживання саджанців і живців, а також для полегшення механізованого догляду за рослинами та ін.
Препарати: ТУР, емістим, метиур, дипромол, фарбізол, ресин, івин, капонин та ін.
Способи застосування синтетичних РРР – намочування насіння, живців у їх розчинах, обприскування вегетуючих органів рослин. Найефективнішим вважається обприскування розчинами фіторегуляторів у поєднанні із засобами хімічного захисту рослин від хвороб і шкідників (так зване застосування бакових сумішей).
Наприклад, виробнича перевірка в господарствах Черкаської, Вінницької, рівненської областей показала, що передпосівна обробка насіння зернових і зернобобових емістимом (1-2 мл/т) разом з мікроелементами (100 мл композиції в 10 л води) або ж обробка вегетуючих рослин: озимих культур весноюпри денній температурі більше 10ºС, ярих зернових і зернобобових у фазі 3-5 листків дали приріст урожаю від 5 до 8,3 ц/га.
Вчені розробляють нетоксичні синтетичні аналоги гормонів шкідників (ОК-37, ОК-84, ОК-85), які одночасно є ефективними засобами захисту рослин. Так, препарат ОК-37 у дозі 4 г/га викликає загибель 90-100% личинок колорадського жука. Ефективні вони також в боротьбі зі шкідниками цукрових буряків (бурякова попелиця, лучний метелик), капусти, проса, картоплі та ін.
З меттою стимуляції бульбоутворення і підвищення вррожайності середньостиглих сортів картоплі рекомендований препарат фарбізол (малотоксичний, використовується шляхом передпосадкового обприскуваннябульб 0,01% водним розчином). Слід зазначити, що цей препарат зменшує вміст нітратів у бульбах, підвищує кількість аскорбінової кислоти.
Для цукрових буряків рекомендований ефективний регулятор росту ресин. Він поліпшує схожість насіння, стійкістьвегетуючих рослиндо коренеїда, підвищує вміст цукру.
3. У рослин розрізняють такі основні типи фітогормонів: ауксини, цитокініни, гібереліни, абсцизова кислота, етилен, брасиностероїди.
Спеціалісти ведуть пошук нових ендогенних фізіологічно активних речовин, які з часом можуть набути статус фітогормонів. Тільки за останні роки до фітогормонів були віднесені жасмонова кислота, олігосахарини, фузікокцин (Муромцев Г.С., 1996). Кожна хімічна категорія фітогормонів має характерний вплив на ріст і спеціалізацію клітин рослин, що використовується ін вітро. Проте треба чітко запам’ятати, що генотип рослин не змінюється під дією будь-якогорегулятора росту. РРР лише допомагають рослинам розкрити їх генетичний потенціалбез появи нових спадкових властивостей. Крім того, РРР ні в якому випадку не замінюють елементів живлення.
Ауксини
Ауксини (від грецького аuxein - збільшуватись, рости) - група фітогормонів, які регулюють процеси поділу та розтягування клітин, сприяють формуванню коренів, провідних пучків, оплодня.
Історія вивчення ауксинів починається з прізвищ Ч.Дарвіна (1880), П.Бойсен-Йєнсена.(1913), Ф.Вента (1926-1928). У роботах цих дослідників доведено, що вигин колеоптиля злаків до однобічного світлового подразника (фототропізм) пов'язаний із синтезом активного хімічного стимулу в їх верхівках і пересуванням його вниз по рослині. Виділення ауксину у чистому вигляді і його ідентифікація були зроблені у 1934 році.
3 хімічної точки зору ця речовина визначається як індоліл-3-оцтова кислота, хоча частіше її називають гетероауксином. Тільки у 1972 році за допомогою мас-пектроскопії було остаточно доведено, що саме індолілоцтова кислота (ЮК) пересувається з верхівок колеоптилів.
ІОК є основним ауксином рослин, але не єдиним. Відомо багато речовин індольної природи з ауксиновою дією, які синтезовані лабораторним шляхом: індолілпоропіонова, індолілмасляна, індолілпіровиноградна, нафтилоцтова кислота. Існують ауксини неіндольної природи: 2,4-дихлорфеноксиоцтова кислота (2,4-Д); 2,4,5-трихлорфеноксиоцтова кислота (2,4,5-Т); 2,3,6-трихлор-бензойна кислота та ін. Вони у високих концентраціях використовуються як гербіциди.

До ауксиноподібних речовин відносять також деякі природні фенольні сполуки: ферулову кислоту, коніфериловий спирт, ванільні, кофейну кислоту. Їх активність нижча, ніж ЮК. Саме те, що знайдено багато сполук із ауксиновою активністю, говорить про ауксин у множині, а не про ауксин як одну хімічну сполуку.
Ауксини утворюються в молодих частинах рослин, які активно ростуть: точках росту стебла, верхівках коренів, молодих листках, бруньках, квітках і плодах. Транспортування ЮК по рослинних тканинах відбувається полярно - від верхівки пагона до кореня, від листка до верхівки пагона або ж вгору по кореню. В тканинах рослин ЮК знаходиться у двох формах: вільний і зв'язаній. Біологічна активність притаманна тільки вільним формам ЮК, Нерівномірним розподілом ауксинів в осьових органах пояснюють ростові рухи, а також тропізми рослин.
Сучасне уявлення про фізіологічну дію ауксинів
Класична дія ауксинів - посилення росту за рахунок стимуляції розтягнення клітин. ЮК активує роботу мембранної помпи іонів Н+, знижує рН позаклітинного середовища і посилює, тим самим, пластичність клітинних стінок. Підвищення концентрації протонів послаблює водневі зв'язки між мікрофібрилами целюлози, геміцелюлози і ксилоглюканів, що дозволяє їм ковзатись відносно одна одної при підвищенні тургорного тиску. Цікавим спостереженням є "кислий ріст" клітин за рахунок розтягнення клітинних стінок без ауксинів при рН близьких до 3,0. Можливо, що у розпушуванні клітинної станки беруть участь ферменти із низькими значеннями рН.
Однак видовження клітини - це не просто розтягнення клітинної стінки. Про це свідчить той факт, що клітинні стінки при розтягненні не стають тоншими, тобто одночасно відбувається синтез de novo компонентів стінки.
Це, в свою чергу, дає підстави враховувати участь ауксинів в синтезі і транспорті нових клітинних полісахаридів. Вченими вже встановлена причетність ІОК до стимуляції синтезу різних типів РНК (м-РНК, т-РНК, р-РНК) і білка в клітинах, які розтягуються.
ІОК зумовлює явище апікального домінування, коли апікальна меристема гальмує ріст бокових меристем. Домінування верхівки — класичний приклад того, як одна частина рослини контролює іншу за допомогою фітогормонів. Природу апікального домінування пояснюють декілька гіпотез:
1) Верхівочна меристема (брунька) найбільш насичена ауксином, і є атрагуючим центром притягнення води та різних поживних речовин, яких не вистачає для бокових бруньок;
2) Під дією ауксину синтезується інгібітор росту, який проникає у бокові меристеми і гальмує їх розвиток.
Синтетичні ауксини широко використовуються на практиці для укорінення черенків, покращення зростання тканин при щепленні, для запобігання опадання зав'язі, а також як сильні гербіциди. Так, суміш 2,4-Д і 2,4,5-Т застосовувалась США під час війни у В'єтнамі (дефоліант "Ейджент Оранж"), Слід зазначити, що ЮК у супероптимальних концентраціях також здатна гальмувати ріст, виступаючи вже інгібітором.
Цитокініни
Цитокініни були відкриті в результаті інтенсивної роботи з культурою тканин, яку проводив американський дослідник Ф.Скуг у 1945-1955 роках. 3 хімічної точки зору вони є похідними 6-амінопурину (аденіну). На сьогодні ідентифіковано близько і5 природних цитокінінів (зеатин, зеатинрибозид, зеатинрибозидфосфат та ін.), В умовах ін вітро широко використовують високоактивний синтетичний препарат кінетин ( одержують при ав-токлавуванні ДНК) і бензиламінопурин /БАП/.
Більше
всього цитокінів там, де проходить
швидкий
поділ клітин. Особливо багато міститься
їх у проростаючому
насінні, достигаючих плодах, верхівках
коренів, пухлинних
клітинах; менше - у стеблах, листках.
Синтез цитокініів у дорослих рослин
локалізується в апікальній меристемі
коренів, звідки вони транспортуються
у надземні органи з ксилемним соком.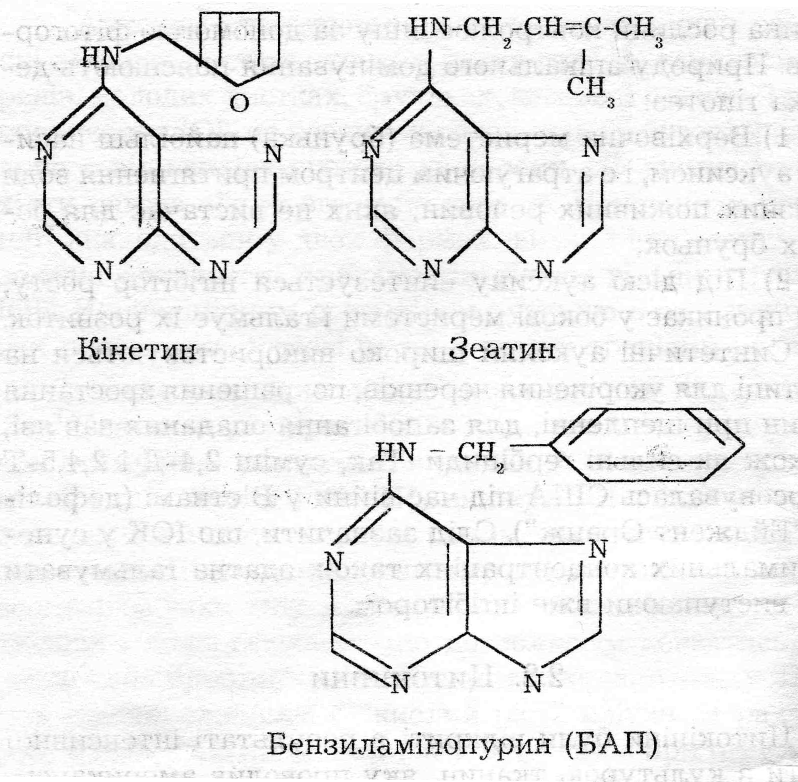
Фізіологічна дія цитокінінів
Цитокініни поліфункціональні у своїй дії на різних етапах росту і розвитку рослин. Сучасна наробка фактичних даних і їх обговорення дозволяють акцентувати увагу на двох основних ефектах фізіологічної дії цитокінінів:
1 - стимуляція процесів клітинного поділу і диференціації;
2 - затримка процесів старіння відокремлених органів.
Структурна подібність цитокінінів з аденіном - компонентом ДНК і РНК - не випадкова і пов'язана з їх функціональними особливостями. Достовірно встановлена специфічність дії цитокінінів як стимуляторів мітозу.
При вирощуванні ізольованих клітин ін вітро особливо чітко виявляється залежність поділу клітин від введення до поживного середовища екзогенного цитокініну.
При взаємодії з ауксинами і гіберелінами цитокініни беруть участь у новоутворенні і диференціації органів ін вітро та ін віво, що пов'язують з їх впливом на біосинтез ДНК, РНК, білків, а також на перерозподіл продуктів обміну у рослині.
Цитокініни стимулюють розвиток латеральних точок росту (бокових бруньок), тобто беруть участь у подоланні апікального домінування. У підрозділі 2.1. ми розглядали, що ауксини утворюються у верхівках, приміщуються вниз по стеблу і пригнічують розвиток бокових бруньок. Чим ближче бокові бруньки розміщені до верхівки, тим сильніше пригнічені, і навпаки. Таке гальмування росту можливо зняти шляхом видалення верхівки або нанесенням цитокініну на бокові бруньки. Саме цим способом домагаються утворення пагонів з усіх пазухових меристем бруньки ін вітро на підвищених концентраціях цитокінінів в поживних середовищах.
Здатність цитокінінів затримувати старіння проявляється на листках. Відокремлений листок у воді швидко старіє, що проявляється у пожовтінні, втраті ним білка, ДНК, РНК. Якщо нанести на такий листок краплю кінетину, то оброблена ділянка залишиться зеленою. Затримка старіння листка також відбувається при утворенні коренів на черешку листка. З цього можна зробити висновок, що цитокініни, які утворюються в коренях, переміщуються в листки і підтримують структурно-функціональну життєдіяльність листків. Цитокініни продовжують тривалість життя ( об'єкти довго зберігають гарний вигляд) свіжої капусти, салату та інших зелених овочів, а також зрізаних квітів, що використовується на практиці.
Гібереліни
Гібереліни були відкриті у 1926 році японським вченим Е.Куросавою при вивченні ураження рису грибом Gibberella. Хворі рослини сильно витягувались у довжину, вилягали і гинули. Екстракт гриба, а також виділена з нього у кристалічному вигляді речовина викликали аналогічну симптоматику захворювання. Ці речовини були названі гіберелінами. (від назви гриба). 3 хімічної точки зору гібереліни відносяться до сполук дитерпеноїдів. Вихідними сполуками для їх синтезу у клітинах є ацетат і мевалонова кислота. Для комерційних цілей гібереліни отримують з культури грибів шляхом мікробіологічного синтезу.
На сьогодні відомо близько 60 природних гіберелінів, які позначаються скорочено ГК1; ГК2 і т.д. Найбільш типовим представником комплексу гіберелінів є гіберелова кислота (ГК3 ), яка знайдена в усіх вивчених рослинах:
У вищих рослин найбільш багаті на гібереліни швидкоростучі органи - молоді апікальні листки, бруньки, незріле насіння і плоди.
Звідси вони мігрують по флоемі і ксилемі уверх-вниз, тобто гібереліни не виявляють полярності транспорту, яка характерна для ауксинів. Велике значення для утворення гіберелінів у рослині має світло (воно сприяє їх біосинтезу в хлоропластах листків).
Фізіологічна дія гіберелінів
Різнопланові експерименти з вивчення фізіологічної дії гіберелінів дозволяють зробити висновок, що гібереліни - компоненти систем, які регулюють ріст і розвиток рослин .
Найбільш виражена дія гіберелінів - їх здатність стимулювати ріст, видовження стебла за рахунок розтягування клітин ( а не їх поділу), що вказує на подібність із ефектом дії ауксинів. За допомогою гіберелінів відновлюють нормальний ріст карликових сортів гороху, кукурудзи або перетворюють карликову форму квасолі у витку ліану. На практиці гіберелінами обробляють коноплю та льон для підвищення виходу волокна.
Другий класичний ефект дії гіберелінів пов'язаний з виходом насіння злакових із стану спокою. Після того, як насіння поглине крізь мікропіле воду, зародок в зоні щитка починає синтезувати гібереліни ГА3, ГА4. Гібереліни переміщуються в алейроновий прошарок, що оточує ендосперм, і стимулюють утворення гідролітичних ферментів (альфа-амілаз та ін.). Ферменти починають розщеплювати запасний крохмаль ендосперму до простих цукрів, які використовуються для росту зародка. Таким чином, гібереліновий "запуск" утворення амілаз є необхідною умовою для проростання насіння. Механізм цього процесу остаточно не з'ясований. Вважають, що гібереліни стимулюють синтез м-РНК, які специфічні для амілаз.
Важливе значення гіберелінів у процесі яровизації і цвітіння. Яровизація - реакція рослин на вплив низьких позитивних температур (+2 - +10° С) у певний період онтогенезу. Центром сприйняття яровизаційного впливу в рослині може бути точка росту або будь-яка зона, в якій відбувається поділ клітин. Яровизація проявляється у прискоренні початку періоду плодоношення (цвітіння). Встановлено, що у ході яровизації підвищується рівень гіберелінів, що дозволяє холодову обробку замінити обробкою неяровизованих рослин гіберелінами. Так, екзогенно введений гіберелін у багатьох дворічних рослин виключає потребу в яровизації і викликає їх цвітіння.