Добавлен: 05.02.2019
Просмотров: 1780
Скачиваний: 21
Из-за низкой температуры осадки выпадают в Антарктике только в виде снега. В такой форме они не могут быть использованы растениями. Вот здесь-то темная окраска лишайников и приходит им на помощь.
Темноокрашенные слоевища антарктических лишайников за счет высокой солнечной радиации быстро нагреваются до положительной температуры даже при отрицательной температуре воздуха. Снег, падающий на эти нагретые слоевища, тает, превращаясь в воду, которую лишайник сразу же впитывает. Таким образом он обеспечивает себя водой, необходимой для осуществления процессов дыхания и фотосинтеза.
Насколько разнообразны слоевища лишайников по окраске, настолько же разнообразны они и по форме. Слоевище может иметь вид корочки, листовидной пластинки или кустика. В зависимости от внешнего облика различают три основных морфологических типа:
На́кипные. Таллом накипных лишайников — это корочка («накипь»), нижняя поверхность плотно срастается с субстратом и не отделяется без значительных повреждений. Это позволяет им жить на крутых склонах гор, деревьях и даже на бетонных стенах. Иногда накипный лишайник развивается внутри субстрата и снаружи совершенно не заметен. Как правило, накипные слоевища небольших размеров, их диаметр составляет всего несколько миллиметров или сантиметров, но иногда может достигать и 20 – 30 см. В природе нередко можно наблюдать, как небольшие по размерам накипные слоевища лишайников, сливаясь друг с другом, образуют на каменистой поверхности скал или стволах деревьев крупные пятна, достигающие в диаметре нескольких десятков сантиметров.
Листоватые. Листоватые лишайники имеют вид пластин разной формы и размера. Они более или менее плотно прикрепляются к субстрату при помощи выростов нижнего коркового слоя. Наиболее простое слоевище листоватых лишайников имеет вид одной крупной округлой листовидной пластинки, достигающей в диаметре 10 – 20 см. Такая пластинка нередко бывает плотной, кожистой, окрашенной в темно серый, темно-коричневый или черный цвет.
Кустистые. По организационному уровню кустистые лишайники представляют высший этап развития слоевища. У кустистых лишайников таллом образует множество округлых или плоских веточек. Растут на земле или свисают с деревьев, древесных остатков, скал. Слоевище кустистых лишайников имеет вид прямостоячего или повисающего кустика, реже неразветвленных прямостоячих выростов. Это позволяет кустистым лишайникам путем изгибов веточек в разные стороны занимать наилучшее положение, при котором водоросли могут максимально использовать свет для осуществления фотосинтеза. Слоевища кустистых лишайников могут быть разных размеров. Высота самых маленьких составляет всего несколько миллиметров, а наиболее крупных 30 – 50 см. Повисающие слоевища кустистых лишайников иногда могут достигать колоссальных размеров.
Внутреннее строение лишайника: коровый слой, гонидиальный слой, сердцевина, нижняя кора, ризоиды. Тело лишайников (таллом) представляет собой переплетение грибных гиф, между которыми находится популяция фотобионта.
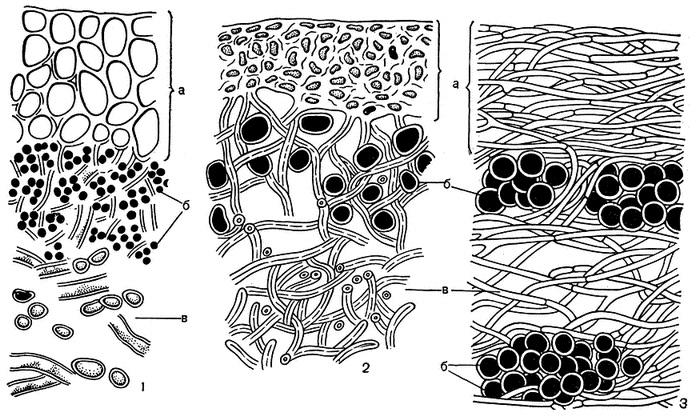
Рис.
2. Анатомическое строение слоевища
лишайников
1 - гетеромерное слоевище (а - верхний коровой слой, б - слой водорослей, в - сердцевина, г - нижний коровой слой); 2 - гомеомерное слоевище слизистого лишайника коллема (Collema flaccidum); 3 - гомеомерное слоевище слизистого лишайника лептогиум (Leptogium saturninum) (а - коровой слой с верхней и нижней стороны слоевища, б - ризоиды)
Каждый из перечисленных анатомических слоев слоевища выполняет в жизни лишайника определенную функцию и в зависимости от этого имеет совершенно определенное строение.
Коровой слой играет в жизни лишайника очень важную роль. Он выполняет сразу две функции: защитную и укрепляющую. Он защищает внутренние слои слоевища от воздействия внешней среды, прежде всего водоросли от чрезмерного освещения. Поэтому коровой слой лишайников обычно бывает плотного строения и окрашен в сероватый, коричневый, оливковый, желтый, оранжевый или красноватый цвет. Коровой слой служит и для укрепления слоевища. Чем выше слоевище поднимается над субстратом, тем более оно нуждается в укреплении. Укрепляющие механические функции в таких случаях нередко выполняет толстый коровой слой. На нижнем коровом слое лишайников обычно образуются органы прикрепления. Иногда они имеют вид очень тонких нитей, состоящих из одного ряда клеток. Эти нити называют ризоидами. Каждая такая нить берет начало от одной клетки нижнего корового слоя. Нередко несколько ризоидов соединяются в толстые ризоидальные тяжи.
В зоне водорослей осуществляются процессы ассимиляции углекислоты и накопление органических веществ. Как известно, для осуществления процессов фотосинтеза водорослям не обходим солнечный свет. Поэтому слой водорослей обычно размещается вблизи верхней поверхности слоевища, непосредственно под верхним коровым слоем, а у вертикально стоящих кустистых лишайников еще и над нижним коровым слоем. Слой водорослей чаще всего бывает небольшой толщины, и водоросли размещаются в нем так, что находятся почти в одинаковых условиях освещения. Водоросли в слоевище лишайника могут образовывать непрерывный слой, но иногда гифы микобионта делят его на отдельные участки. Для осуществления процессов ассимиляции углекислоты и дыхания водорослям необходим также нормальный газообмен. Поэтому грибные гифы в зоне водорослей не образуют плотных сплетений, а расположены рыхло на некотором расстоянии друг от друга.
Под слоем водорослей расположен сердцевинный слой. Обычно сердцевина по толщине значительно превышает коровой слой и зону водорослей. От степени развития сердцевины зависит толщина самого слоевища. Основная функция сердцевинного слоя – проведение воздуха к клеткам водорослей, содержащим хлорофилл. Поэтому для большинства лишайников характерно рыхлое расположение гиф в сердцевине. Воздух, попадающий в слоевище, легко проникает к водорослям по промежуткам между гифами. Сердцевинные гифы слабоветвисты, с редкими поперечными перегородками, с гладкими, слабожелатинообразными толстыми стенками и довольно узким просветом, заполненным протоплазмой. У большинства лишайников сердцевина белая, так как гифы сердцевинного слоя бесцветны.
По внутреннему строению лишайники разделяют на:
— гомеомерные (Collema), клетки фотобионта распределены хаотично среди гиф гриба по всей толщине таллома;
— гетеромерные (Peltigera canina), таллом на поперечном срезе можно чётко разделить на слои.
Лишайников с гетеромерным талломом большинство. В гетеромерном талломе верхний слой — корковый, сложенный гифами гриба. Он защищает таллом от высыхания и механических воздействий. Следующий от поверхности слой — гонидиальный, или альгальный, в нём располагается фотобионт. В центре располагается сердцевина, состоящая из беспорядочно переплетенных гиф гриба. В сердцевине в основном запасается влага, она также играет роль скелета. У нижней поверхности таллома часто находится нижняя кора, с помощью выростов которой (ризин) лишайник прикрепляется к субстрату. Полный набор слоёв встречается не у всех лишайников.
Как и в случае двухкомпонентных лишайников, водорослевый компонент — фикобионт — трёхкомпонентных лишайников равномерно распределен по таллому, либо образует слой под верхней корой. Некоторые трёхкомпонентные цианолишайники образуют специализированные поверхностные или внутренние компактные структуры (цефалодии), в которых сосредоточен цианобактериальный компонент.
Способы питания лишайников
Лишайники представляют для физиологических исследований сложный объект, так как состоят из двух физиологически противоположных компонентов – гетеротрофного гриба и автотрофной водоросли. Поэтому приходится сначала отдельно изучать жизнедеятельность мико- и фикобионта, что делается с помощью культур, а затем жизнь лишайника как целостного организма. Понятно, что такая «тройная физиология» – трудный путь исследования, и не удивительно, что в жизнедеятельности лишайников кроется еще много загадочного. Однако общие закономерности их обмена веществ все же выяснены.
Довольно много исследований посвящено процессу фотосинтеза у лишайников. Так как лишь небольшая часть их слоевища (5 – 10% объема) образована водорослью, которая тем не менее является единственным источником снабжения органическими веществами, встает существенный вопрос об интенсивности фотосинтеза в лишайниках.
Как показали измерения, интенсивность фотосинтеза у лишайников намного ниже, чем у высших автотрофных растений.
Для нормальной фотосинтетической активности слоевище должно содержать определенное количество воды, зависящее от анатомоморфологического типа лишайника. В общем в толстых слоевищах оптимальное содержание воды для активного фотосинтеза ниже, чем в тонких и рыхлых слоевищах. При этом весьма существенно то обстоятельство, что многие виды лишайников, особенно в сухих местообитаниях, вообще редко или по крайней мере очень нерегулярно снабжаются оптимальным количеством внутрислоевищной воды. Ведь регуляция водного режима у лишайников происходит совсем по-иному, чем у высших растений, имеющих специальный аппарат, способный контролировать получение и расходование воды. Лишайники усваивают воду (в виде дождя, снега, тумана, росы и пр.) очень быстро, но пассивно всей поверхностью своего тела и отчасти ризоидами нижней стороны. Такое поглощение слоевищем воды представляет собой простой физический процесс, как, например, впитывание воды фильтровальной бумагой. Лишайники способны впитывать воду в очень больших количествах, обычно до 100 – 300% от сухой массы слоевища, а некоторые слизистые лишайники (коллемы, лептогиумы и др.) даже до 800 – 3900%.
Минимальное содержание воды в лишайниках в природных условиях составляет примерно 2 – 15% от сухой массы слоевища.
Отдача воды слоевищем также происходит довольно быстро. Насыщенные водой лишайники на солнце через 30 – 60 мин теряют всю свою воду и делаются хрупкими, т. е. содержание воды в слоевище становится ниже минимально необходимого для активного фотосинтеза. Из этого вытекает своеобразная «аритмичность» фотосинтеза лишайников – его продуктивность меняется в течение дня, времени года, ряда годов в зависимости от общих экологических условий, особенно гидрологических и температурных.
Имеются наблюдения, что многие лишайники более активно фотосинтезируют в утренние и вечерние часы и что фотосинтез продолжается у них и зимой, а у напочвенных форм даже под нетолстым снеговым покровом.
Важным компонентом в питании лишайников является азот. Те лишайники, которые имеют в качестве фикобионта зеленые водоросли (а их большинство), воспринимают соединения азота из водных растворов, когда их слоевища пропитываются водой. Возможно, что часть азотистых соединений лишайники берут и прямо из субстрата – почвы, коры деревьев и т. д. Экологически интересную группу составляют так называемые нитрофильные лишайники, растущие в местообитаниях, богатых азотистыми соединениями, – на «птичьих камнях», где много экскрементов птиц, на стволах деревьев и т. д. (виды ксантории, фисции, калоплаки и др.). Лишайники, имеющие в качестве фикобионта сине-зеленые водоросли (особенно ностоки), способны фиксировать атмосферный азот, так как этой способностью обладают содержащиеся в них водоросли. В опытах с такими видами (из родов коллема, лептогиум, пельтигера, лобария, стикта и др.) было установлено, что их слоевища быстро и активно поглощают атмосферный азот. Эти лишайники часто селятся на субстратах, весьма бедных азотистыми соединениями. Большая часть азота, фиксированного водорослью, направляется микобионту и лишь незначительная часть используется самим фикобионтом. Имеются данные, что микобионт в слоевище лишайника ведет активный контроль над освоением и распределением азотистых соединений, фиксированных из атмосферы фикобионтом.
Описанный выше ритм жизни является одной из причин для очень медленного роста большинства лишайников. Иногда лишайники растут всего лишь на несколько десятых миллиметра в год, в основном менее чем на один сантиметр. Другой причиной медленного роста является то, что фотобионт, составляя нередко менее 10% объёма лишайника, берёт на себя обеспечение микобионта питательными веществами. В хороших условиях, с оптимальными влажностью и температурой, например в туманных или дождливых тропических лесах, лишайники растут на несколько сантиметров в год.
Ростовая зона лишайников у накипных форм находится по краю лишайника, у листоватых и кустистых на каждой верхушке.
Лишайники являются одними из самых долгоживущих организмов и могут достигать возраста нескольких сотен лет, а в некоторых случаях более 4500 лет, как например Rhizocaгрon geographicum, живущий в Гренландии.
Размножение лишайников
Лишайники размножаются либо спорами, которые образует микобионт половым или бесполым путем, либо вегетативно – фрагментами слоевища, соредиями и изидиями.
При половом размножении на слоевищах лишайников в результате полового процесса формируются половые спороношения в виде плодовых тел. Среди плодовых тел у лишайников различают апотеции, перитеции и гастеротеции. Большинство лишайников формируют открытые плодовые тела в виде апотециев – дисковидных образований. Некоторые имеют плодовые тела в форме перитеция – закрытого плодового тела, имеющего вид маленького кувшина с отверстием наверху. Небольшое количество лишайников образуют узкие плодовые тела удлиненной формы, которые называют гастеротециями.
В апотециях, перитециях и гастеротециях споры развиваются внутри сумок – особых мешковидных образований. Лишайники, формирующие споры в сумках, объединяются в большую группу сумчатых лишайников. Они произошли от грибов класса аскомицетов и представляют основную эволюционную линию развития лишайников.
У небольшой группы лишайников споры образуются не внутри сумок, а экзогенно, на вершине удлиненно-булавовидных гиф – базидий, на концах которых развиваются четыре споры. Лишайники с таким образованием спор объединяются в группу базидиальных лишайников.
Женский половой орган лишайников – архикарп – состоит из двух частей. Нижняя часть носит название аскогона и представляет собой спирально закрученную гифу, более толстую по сравнению с другими гифами и состоящую из 10 – 12 одно- или много ядерных клеток. От аскогона вверх отходит трихогина – тоненькая вытянутая гифа, которая проходит через зону водорослей и коровой слой и выходит на поверхность слоевища, возвышаясь над ней своей липкой верхушкой.
Развитие и созревание плодового тела у лишайников – очень медленный процесс, который длится 4 – 10 лет. Сформировавшееся плодовое тело тоже является многолетним, способным в течение ряда лет продуцировать споры. Сколько же спор способны продуцировать плодовые тела лишайников? Подсчитано, например, что у лишайника солорина в апотеции диаметром 5 мм образуется 31 тыс. сумок, а в каждой сумке обычно развивается по 4 споры. Следовательно, общее количество спор, продуцируемое одним апотецием, равно 124 000. В течение одного дня из такого апотеция выбрасывается от 1200 до 1700 спор. Конечно, не все выброшенные из плодового тела споры прорастают. Многие из них, попав в неблагоприятные условия, погибают. Для прорастания споры необходимы прежде всего достаточная влажность и определенная температура.