ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 05.07.2024
Просмотров: 164
Скачиваний: 0
vsdma.com
|
№ 1 Место микробиологии и иммунологии в современной медицине. Роль микробиологии и иммунологии в подготовке врачей-клиницистов и врачей профилактической службы.
Микробиология — наука, изучающая строение, жизнедеятельность и экологию микроорганизмов — мельчайших форм жизни растительного или животного происхождения, не видимых невооруженным глазом. Микробиология изучает всех представителей микромира (бактерии, грибы, простейшие, вирусы). По своей сути микробиология является биологической фундаментальной наукой. Для изучения микроорганизмов она использует методы других наук, прежде всего физики, биологии, биоорганической химии, молекулярной биологии, генетики, цитологии, иммунологии. Как и всякая наука, микробиология подразделяется на общую и частную. Общая микробиология изучает закономерности строения и жизнедеятельности микроорганизмов на всех уровнях — молекулярном, клеточном, популяционном; генетику и взаимоотношения их с окружающей средой. Предметом изучения частной микробиологии являются отдельные представители микромира в зависимости от проявления и влияния их на окружающую среду, живую природу, в том числе человека. К частным разделам микробиологии относятся: медицинская, ветеринарная, сельскохозяйственная, техническая (раздел биотехнологии), морская, космическая микробиология. Многочисленные открытия в области микробиологии, изучение взаимоотношений между макро- и микроорганизмами во второй половине XIX в. способствовали началу бурного развития иммунологии. Вначале иммунология рассматривалась как наука о невосприимчивости организма к инфекционным болезням. В настоящее время она стала общемедицинской и общебиологической наукой. Доказано, что иммунная система служит для защиты организма не только от микробных агентов, но и от любых генетически чужеродных организму веществ с целью сохранения постоянства внутренней среды организма, т.е. гомеостаза. Иммунология является основой для разработки лабораторных методов диагностики, профилактики и лечения инфекционных и многих неинфекционных болезней, а также разработки иммунобиологических препаратов (вакцин, иммуноглобулинов, иммуномодуляторов, аллергенов, диагностических препаратов). Разработкой и производством иммунобиологических препаратов занимается иммунобиотехнология — самостоятельный раздел иммунологии. Современная медицинская микробиология и иммунология достигли больших успехов и играют огромную роль в диагностике, профилактике и лечении инфекционных и многих не инфекционных болезней, связанных с нарушением иммунной системы (онкологические, аутоиммунные болезни, трансплантация органов и тканей и др.). |
№ 2 Основные этапы развития микробиологии и иммунологии. Работы Л. Пастера, Р. Коха и их значение для развития микробиологии и иммунологии.
Основные этапы развития микробиологии и имунологии. Историю развития микробиологии можно разделить на пять этапов: эвристический, морфологический, физиологический, иммунологический и молекулярно-генетический. Пастер сделал ряд выдающихся открытий. За короткий период с 1857 по 1885 г. он доказал, что брожение (молочнокислое, спиртовое, уксуснокислое) не является химическим процессом, а его вызывают микроорганизмы; опроверг теорию самозарождения; открыл явление анаэробиоза, т.е. возможность жизни микроорганизмов в отсутствие кислорода; заложил основы дезинфекции, асептики и антисептики; открыл способ предохранения от инфекционных болезней с помощью вакцинации. Многие открытия Л. Пастера принесли человечеству огромную практическую пользу. Путем прогревания (пастеризации) были побеждены болезни пива и вина, молочнокислых продуктов, вызываемые микроорганизмами; для предупреждения гнойных осложнений ран введена антисептика; на основе принципов Л. Пастера разработаны многие вакцины для борьбы с инфекционными болезнями. Однако значение трудов Л. Пастера выходит далеко за рамки только этих практических достижений. Л. Пастер вывел микробиологию и иммунологию на принципиально новые позиции, показал роль микроорганизмов в жизни людей, экономике, промышленности, инфекционной патологии, заложил принципы, по которым развиваются микробиология и иммунология и в наше время. Л. Пастер был, кроме того, выдающимся учителем и организатором науки. Работы Л. Пастера по вакцинации открыли новый этап в развитии микробиологии, по праву получивший название иммунологического. Принцип аттенуации (ослабления) микроорганизмов с помощью пассажей через восприимчивое животное или при выдерживании микроорганизмов в неблагоприятных условиях (температура, высушивание) позволил Л. Пастеру получить вакцины против бешенства, сибирской язвы, куриной холеры; этот принцип до настоящего времени используется при приготовлении вакцин. Следовательно, Л. Пастер является основоположником научной иммунологии, хотя и до него был известен метод предупреждения оспы путем заражения людей коровьей оспой, разработанный английским врачом Э. Дженнером. Однако этот метод не был распространен на профилактику других болезней. Роберт Кох. Физиологический период в развитии микробиологии связан также с именем немецкого ученого Роберта Коха, которому принадлежит разработка методов получения чистых культур бактерий, окраски бактерий при микроскопии, микрофотографии. Известна также сформулированная Р. Кохом триада Коха, которой до сих пор пользуются при установлении возбудителя болезни. |
№ 3 Роль И. И. Мечникова в формировании учения об иммунитете. Значение открытия Д. И. Ивановского. Роль отечественных ученых (И. Ф. Гамалея, П. Ф. Здродовский, А. А. Смородинцев, М. П. Чумаков, 3. В. Ермольева, В. М. Жданов и др.) в развитии микробиологии и вирусологии.
После работ Л. Пастера появилось множество исследований, в которых пытались объяснить причины и механизмы формирования иммунитета после вакцинации. Выдающуюся роль в этом сыграли работы И. И. Мечникова и П. Эрлиха. Исследования И. И. Мечникова (1845—1916) показали, что большую роль в формировании иммунитета играют особые клетки — макро- и микрофаги. Эти клетки поглощают и переваривают чужеродные частицы, в том числе бактерии. Исследования И. И. Мечникова по фагоцитозу убедительно доказали, что, помимо гуморального, существует клеточный иммунитет. И. И. Мечников, ближайший помощник и последователь Л. Пастера, заслуженно считается одним из основоположников иммунологии. Его работы положили начало изучению иммунокомпетентных клеток как морфологической основы иммунной системы, ее единства и биологической сущности. Д.И.Ивановский (1864— 1920) открыл вирусы — представителей царства vira. Один из основоположников вирусологии. Впервые открыл проходящий через бактериологические фильтры возбудитель табачной мозаики, названный впоследствии вирусом. Труды по фитопатологии и физиологии растений. Гамалея - выдающийся микробиолог. Вместе с И. И. Мечниковым в 1886 году организовал в Одессе первую в России бактериологическую станцию. Автор многих работ по микробиологии и иммунологии (по профилактике холеры, чумы, оспы, паразитарных тифов, бешенства). Открыл бактериолизины, возбудители холеры птиц. Обосновал значение дезинсекции для ликвидации сыпного и возвратного тифов. В 1888 году ученый издал книгу "О прививках против сибирской язвы". Здровский (1890-1976 года), российский микробиолог, иммунолог и эпидемиолог, академик АМН. Исследования по проблемам тропических болезней, бруцеллеза и др. Под руководством Здродовского разработаны методы вакцинации против столбняка, дифтерии и др. инфекций. Автор книги "Учение о риккетсиях и риккетсиозах" Смородинцев, российский вирусолог и иммунолог. Труды по этиологии и профилактике гриппа, энцефалитов и др. вирусных инфекций. Совместно с М. П. Чумаковым разработал и внедрил вакцину против полиомиелита. Ермольева, российский микробиолог. Получила первые отечественные образцы антибиотиков - пенициллина, стрептомицина и др.; интерферона. Жданов, российский вирусолог. Труды по вирусным инфекциям, молекулярной биологии и классификации вирусов, эволюции инфекционных болезней. |
|
№ 4 Основные принципы классификации микробов.
Микробы, или микроорганизмы (бактерии, грибы, простейшие, вирусы), систематизированы по их сходству, различиям и взаимоотношениям между собой. Этим занимается специальная наука — систематика микроорганизмов. Систематика включает три части: классификацию, таксономию и идентификацию. В основу таксономии микроорганизмов положены их морфологические, физиологические, биохимические и молекулярно-биологические свойства. Различают следующие таксономические категории: царство, подцарство, отдел, класс, порядок, семейство, род, вид, подвид и др. В рамках той или иной таксономической категории выделяют таксоны — группы организмов, объединенные по определенным однородным свойствам. Микроорганизмы представлены доклеточными формами (вирусы — царство Vira) и клеточными формами (бактерии, архебактерии, грибы и простейшие). Различают 3 домена (или «империи»): «Bacteria», «Archaea» и «Eukarya»: □ домен «Bacteria» — прокариоты, представленные настоящими бактериями (эубактериями); □ домен «Archaea» — прокариоты, представленные архебактериями; □ домен «Eukarya» — эукариоты, клетки которых имеют ядро с ядерной оболочкой и ядрышком, а цитоплазма состоит из высокоорганизованных органелл — митохондрий, аппарата Гольджи и др. Домен «Eukarya» включает: царство Fungi (грибы); царство животных Animalia (включает прстейшие – подцарство Protozoa); царство растений Plante. Домены включают царства, типы, классы, порядки, семейства, роды, виды. Вид. Одной из основных таксономических категорий является вид (species). Вид — это совокупность особей, объединенных по близким свойствам, но отличающихся от других представителей рода. Чистая культура. Совокупность однородных микроорганизмов, выделенных на питательной среде, характеризующихся сходными морфологическими, тинкториальными (отношение к красителям), культуральными, биохимическими и антигенными свойствами, называется чистой культурой. Штамм. Чистая культура микроорганизмов, выделенных из определенного источника и отличающихся от других представителей вида, называется штаммом. Штамм — более узкое понятие, чем вид или подвид. Клон. Близким к понятию штамма является понятие клона. Клон представляет собой совокупность потомков, выращенных из единственной микробной клетки. Для обозначения некоторых совокупностей микроорганизмов, отличающихся по тем или иным свойствам, употребляется суффикс var (разновидность) вместо ранее применявшегося type. |
№ 5 Принципы классификации бактерий.
Для бактерий рекомендованы следующие таксономические категории: класс, отдел, порядок, семейство, род, вид. Название вида соответствует бинарной номенклатуре, т. е. состоит из двух слов. Например, возбудитель сифилиса пишется как Treponema pallidum. Первое слово — название рода и пишется с прописной буквы, второе слово обозначает вид и пишется со строчной буквы. При повторном упоминании вида родовое название сокращается до начальной буквы, например: Т. pallidum. Бактерии относятся к прокариотам, т. е. доядерным организмам, поскольку у них имеется примитивное ядро без оболочки, ядрышка, гистонов, а в цитоплазме отсутствуют высокоорганизованные органеллы (митохондрии, аппарат Гольджи, лизосомы и др.). Бактерии делят на 2 домена: «Bacteria» и «Archaea». В домене «Bacteria» можно выделить следующие бактерии: 1) бактерии с тонкой клеточной стенкой, грамотрицательные; 2) бактерии с толстой клеточной стенкой, грамположительные; 3) бактерии без клеточной стенки (класс Mollicutes — микоплазмы) Архебактерии не содержат пептидогликан в клеточной стенке. Они имеют особые рибосомы и рибосомные РНК (рРНК). Среди тонкостенных грамотрицательных эубактерий различают: • сферические формы, или кокки (гонококки, менингококки, вейлонеллы); • извитые формы — спирохеты и спириллы; • палочковидные формы, включая риккетсии. К толстостенным грамположительным эубактериям относят: • сферические формы, или кокки (стафилококки, стрептококки, пневмококки); • палочковидные формы, а также актиномицеты (ветвящиеся, нитевидные бактерии), коринебактерии (булавовидные бактерии), микобактерии и бифидобактерии. Тонкостенные грамотрицательные бактерии: Менингококки, гонококки, Вейлонеллы, Палочки, Вибрионы, Кампилобактерии, Хеликобактерии, Спириллы, Спирохеты, Риккетсии, Хламидии. Толстостенные грамположительные бактерии: Пневмококки, Стрептококки, Стафилококки, Палочки, Бациллы, Клостридии, Коринебактерии, Микобактерии, Бифидобактерии, Актиномицеты. |
№ 6 Принципы классификации грибов.
Грибы относятся к царству Fungi (Mycetes, Mycota). Это многоклеточные или одноклеточные нефотосинтезирующие (бес-хлорофильные) эукариотические микроорганизмы с клеточной стенкой. Классификация грибов. Грибы можно разделить на 7 классов: хитридиомицеты, гифохитридиомицеты, оомицеты, зигомицеты, аскомицеты, базидиомицеты, дейтеромицеты. Среди фикомицетов различают: хитридиомицеты, или водные грибы, ведущие сапрофитический образ жизни или поражающие водоросли; гифохитридиомицеты, имеющие сходство с хитридиомицетами и оомицетами; оомицеты — паразиты высших растений и водяные плесени; зигомицеты включают представителей рода Mucor, распространенных в почве и воздухе и способных (например, грибы рода Mucor) вызывать мукоромикоз легких, головного мозга и других органов. При бесполом размножении на плодоносящей гифеспорангиеносце образуется спорангий — шаровидное утолщение с оболочкой, содержащее многочисленные споры (спорангиоспоры). Половое размножение (оогамия) у зигомицетов осуществляются путем образования зигоспор, или ооспор. Эумицеты представлены аскомицетами и базидиомицетами (совершенные грибы), а также дейтеромицетами (несовершенные грибы). Аскомицеты (или сумчатые грибы) объединяют группу грибов, имеющих септированный мицелий и отличающихся способностью к половому размножению. Свое название аскомицеты получили от основного органа плодоношения — сумки, или аска, содержащего 4 или 8 гаплоидных половых спор (аскоспор). К аскомицетам относятся представители родов Aspergillus, Penicillium и др., отличающиеся особенностями формирования плодоносящих гиф. У Aspergillus (леечная плесень) на концах плодоносящих гифконидиеносцев имеются утолщения — стеригмы, на которых образуются цепочки спор — конидии. Некоторые виды аспергилл могут вызывать аспергиллезы и афлатоксикозы. Плодоносящая гифа у грибов рода Penicillium (кистевик) напоминает кисточку, так как из нее (на конидиеносце) образуются утолщения, разветвляющиеся на более мелкие структуры — стеригмы, на которых находятся цепочки конидий. Пеницициллы могут вызывать заболевания (пенициллинозы). Многие виды аскомицетов являются продуцентами антибиотиков. Представителями аскомицетов являются и дрожжи — одноклеточные грибы, утратившие способность к образованию истинного мицелия. Дрожжи имеют овальную форму клеток, диаметр которых 3—15 мкм. Они размножаются почкованием, бинарным делением (делятся на две равные клетки) или половым путем с образованием аскоспор. Дрожжи используют в биотехнологических процессах. Заболевания, вызываемые некоторыми видами дрожжей, получили название дрожжевых микозов. К аскомицетам относится и возбудитель эрготизма, или спорыньи (Claviceps purpurea), паразитирующий на злаках. Базидиомицеты — шляпочные грибы с септированным мицелием. Дейтеромицеты — несовершенные грибы (Fungi imperfecti) — являются условным классом грибов, объединяющим грибы с септированным мицелием, не имеющих полового размножения. Они размножаются только бесполым путем, образуя конидии. К несовершенным грибам относятся грибы рода Candida, поражающие кожу, слизистые оболочки и внутренние органы (кандидоз). Они имеют овальную форму, диаметр 2—5 мкм; делятся почкованием (бластоспоры), образуют псевдомицелий (почкующиеся клетки из ростковой трубочки вытягиваются в нить), на концах которого находятся хламидоспоры. Эти грибы называют дрожжеподобными. Истинные дрожжи (аскомицеты) образуют аскоспоры, не имеют псевдомицелия и хламидоспор. Подавляющее большинство грибов, вызывающих заболевания у человека (микозы), относятся к несовершенным грибам. |
|
|
||
|
№ 7 Принципы классификации простейших.
Простейшие представлены 7 типами, из которых четыре типа (Sarcomastigophora, Apicomplexa, Ciliopkora, Microspora) включают возбудителей заболеваний у человека. Тип Sarcomastigophora. Подтип Mastigophora (жгутиконосцы) включает следующих патогенных представителей: трипаносому — возбудителя африканского трипаносомоза (сонная болезнь); лейшмании — возбудителей кожной и висцеральной форм лейшманиозов; трихомонады, передающиеся половым путем и паразитирующие в толстой кишке человека; лямблию — возбудителя лямблиоза. Эти простейшие характеризуются наличием жгутиков: один — у лейшмании, четыре свободных жгутика и короткая ундулирующая мембрана — у трихомонад. К подтипу Sarcodina (саркодовые) относится дизентерийная амеба — возбудитель амебной дизентерии человека. Морфологически сходна с ней непатогенная кишечная амеба. Эти простейшие передвигаются путем образования псевдоподий. Питательные вещества захватываются и погружаются в цитоплазму клеток. Половой путь размножения у амеб отсутствует. При неблагоприятных условиях они образуют цисту. Тип Apicomplexa. В классе Sporozoa (споровики) патогенными представителями являются возбудители токсоплазмоза, кокцидиоза, саркоцистоза и малярии. Жизненный цикл возбудителей малярии характеризуется чередованием полового размножения (в организме комаров Anopheles) и бесполого (в клетках тканей и эритроцитах человека они размножаются путем множественного деления). Токсоплазмы имеют форму полулуний. Токсоплазмозом человек заражается от животных. Токсоплазмы могут передаваться через плаценту и поражать центральную нервную систему и глаза плода. Тип Ciliophora. Патогенный представитель — возбудитель балантидиаза — поражает толстый кишечник человека. Балантидии имеют многочисленные реснички и поэтому подвижны. Тип Microspora включает микроспоридии — маленькие (0,5—10 мкм) облигатные внутриклеточные паразиты, широко распространенные среди животных и вызывающие у ослабленных людей диарею и гнойно-воспалительные заболевания. |
№ 8 Принципы классификации вирусов.
В основу классификации вирусов положены следующие категории: • тип нуклеиновой кислоты (ДНК или РНК), ее структура, количество нитей (одна или две), особенности воспроизводства вирусного генома; • размер и морфология вирионов, количество капсомеров и тип симметрии; • наличие суперкапсида; • чувствительность к эфиру и дезоксихолату; • место размножения в клетке; • антигенные свойства и пр. Вирусы имеют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК-содержащие вирусы. Они обычно гаплоидны, т.е. имеют один набор генов. Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными. Среди РНК- содержащих вирусов различают вирусы с положительным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК). Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию. Вирусы — мельчайшие микробы, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только ДНК или РНК. Относятся к царству Vira. Являясь облигатными внутриклеточными паразитами, вирусы размножаются в цитоплазме или ядре клетки. Они — автономные генетические структуры. Отличаются особым — разобщенным (дисъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки, затем происходит их сборка в вирусные частицы. Сформированная вирусная частица называется вирионом. Морфологию вирусов изучают с помощью электронной микроскопии, так как их размеры малы (18-400 нм) и сравнимы с толщиной оболочки бактерий. Форма вирионов может быть различной: палочковидной (вирус табачной мозаики), пулевидной (вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), нитевидной (филовирусы), в виде сперматозоида (многие бактериофаги). Различают просто устроенные и сложно устроенные вирусы. Простые, или безоболочечные, вирусы состоят из нуклеиновой кислоты и белковой оболочки, называемой капсидом. Капсид состоит из повторяющихся морфологических субъединиц — капсомеров. Нуклеиновая кислота и капсид взаимодействуют друг с другом, образуя нуклеокапсид. Сложные, или оболочечные, вирусы снаружи капсида окружены ли-попротеиновой оболочкой (суперкапсидом, или пеплосом). Эта оболочка является производной структурой от мембран вирус-инфицированной клетки. На оболочке вируса расположены гликопротеиновые шипы, или шипики (пепломеры). Под оболочкой некоторых вирусов находится матриксный М-белок. Тип симметрии. Капсид или нуклеокапсид могут иметь спиральный, икосаэдрический (кубический) или сложный тип симметрии. Икосаэдрический тип симметрии обусловлен образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту (например, у вирусов гепатита А, герпеса, полиомиелита). Спиральный тип симметрии обусловлен винтообразной структурой нуклеокапсида (например, у вируса гриппа). |
|
|
|
||
|
|
||
|
№ 9 Морфологические и тинкториальные свойства бактерий. Методы окраски.
Морфологические свойства бактерий. Бактерии — микроорганизмы, не имеющие оформленного ядра (прокариоты). Бактерии имеют разнообразную форму и довольно сложную структуру, определяющую многообразие их функциональной деятельности. Для бактерий характерны четыре основные формы: сферическая (шаровидная), цилиндрическая (палочковидная), извитая и нитевидная. Бактерии шаровидной формы — кокки — в зависимости от плоскости деления и расположения относительно друг друга отдельных особей подразделяются на микрококки (отдельно лежащие кокки), диплококки (парные кокки), стрептококки (цепочки кокков), стафилококки (имеющие вид виноградных гроздьев), тетракокки (образования из четырех кокков) и сарцины (пакеты из 8 или 16 кокков). Палочковидные бактерии располагаются в виде одиночных клеток, дипло- или стрептобактерий. Извитые формы бактерий — вибрионы и спириллы, а также спирохеты. Вибрионы имеют вид слегка изогнутых палочек, спириллы — извитую форму с несколькими спиральными завитками. Размеры бактерий колеблются от 0,1 до 10 мкм. В состав бактериальной клетки входят капсула, клеточная стенка, цитоплаз-матическая мембрана и цитоплазма, в которой содержатся нук-леоид, рибосомы и включения. Некоторые бактерии снабжены жгутиками и ворсинками. Ряд бактерий образуют споры, которые располагаются терминально, субтерминально или центрально; превышая поперечный размер клетки, споры придают ей веретенообразную форму. Методы окраски. Окраску мазка производят простыми или сложными методами. Простые заключаются в окраске препарата одним красителем; сложные методы (по Граму, Цилю — Нильсену и др.) включают последовательное использование нескольких красителей и имеют дифференциально-диагностическое значение. Отношение микроорганизмов к красителям расценивают как тинкториальные свойства. Существуют специальные методы окраски, которые используют для выявления жгутиков, клеточной стенки, нуклеоида и разных цитоплазматических включений. При простых методах мазок окрашивают каким-либо одним красителем, используя красители анилинового ряда (основные или кислые). Если красящий ион (хромофор) — катион, то краситель обладает основными свойствами, если хромофор - анион, то краситель имеет кислые свойства. Кислые красители — эритрозин, кислый фуксин, эозин. Основные красители — генциановый фиолетовый, кристаллический фиолетовый, метиленовый синий, основной фуксин. Преимущественно для окраски микроорганизмов используют основные красители, которые более интенсивно связываются кислыми компонентами клетки. Из сухих красителей, продающихся в виде порошков, готовят насыщенные спиртовые растворы, а из них — водно-спиртовые, которые и служат для окрашивания микробных клеток. Микроорганизмы окрашивают, наливая краситель на поверхность мазка на определенное время. Окраску основным фуксином ведут в течение 2 мин, метиленовым синим — 5—7 мин. Затем мазок промывают водой до тех пор, пока стекающие струи воды не станут бесцветными, высушивают осторожным промоканием фильтровальной бумагой и микроскопируют в иммерсионной системе. Если мазок правильно окрашен и промыт, то поле зрения совершенно прозрачно, а клетки интенсивно окрашены. Сложные методы окраски применяют для изучения структуры клетки и дифференциации микроорганизмов. Окрашенные мазки микроскопируют в иммерсионной системе. Последовательно нанести на препарат определенные красители, различающиеся по химическому составу и цвету, протравы, спирты, кислоту и др. Существуют несколько основных окрасок: по Граму, по Цилю-Нельсону, по Ауески, Нейссера, Бури-Гинса. |
№ 10 Структура и химический состав бактериальной клетки. Особенности строения грамположительных и грамотрицательных бактерий.
Бактериальная клетка состоит из клеточной стенки, цитоплазматической мембраны, цитоплазмы с включениями и ядра, называемого нуклеоидом. Имеются дополнительные структуры: капсула, микрокапсула, слизь, жгутики, пили. Некоторые бактерии в неблагоприятных условиях способны образовывать споры. Клеточная стенка. В клеточной стенке грамположительных бактерий содержится небольшое количество полисахаридов, липидов, белков. Основным компонентом толстой клеточной стенки этих бактерий является многослойный пептидогликан (муреин, мукопептид), составляющий 40-90 % массы клеточной стенки. С пептидогликаном клеточной стенки грамположительных бактерий ковалентно связаны тейхоевые кислоты (от греч. teichos — стенка). В состав клеточной стенки грамотрицательных бактерий входит наружная мембрана, связанная посредством липопротеина с подлежащим слоем пептидогликана. На ультратонких срезах бактерий наружная мембрана имеет вид волнообразной трехслойной структуры, сходной с внутренней мембраной, которую называют цитоплазматической. Основным компонентом этих мембран является бимолекулярный (двойной) слой липидов. Внутренний слой наружной мембраны представлен фосфолипидами, а в наружном слое расположен липополисахарид. Функции клеточной стенки: 1. Обусловливает форму клетки. 2. Защищает клетку от механических повреждений извне и выдерживает значительное внутреннее давление. 3. Обладает свойством полупроницаемости, поэтому через нее избирательно проникают из среды питательные вещества. 4. Несет на своей поверхности рецепторы для бактериофагов и различных химических веществ. Метод выявления клеточной стенки - электронная микроскопия, плазмолиз. L-формы бактерий, их медицинское значение L-формы - это бактерии, полностью или частично лишенные клеточной стенки (протопласт +/- остаток клеточной стенки), поэтому имеют своеобразную морфологию в виде крупных и мелких сферических клеток. Способны к размножению. Цитоплазматическая мембрана располагается под клеточной стенкой (между ними - периплазматическое пространство). По строению является сложным липидобелковым комплексом, таким же, как у клеток эукариот (универсальная мембрана). Функции цитоплазматической мембраны: 1. Является основным осмотическим и онкотическим барьером. 2. Участвует в энергетическом метаболизме и в активном транспорте питательных веществ в клетку, так как является местом локализации пермеаз и ферментов окислительного фосфорилирования. 3. Участвует в процессах дыхания и деления. 4. Участвует в синтезе компонентов клеточной клетки (пептидогликана). 5. Участвует в выделении из клетки токсинов и ферментов. Цитоплазматическая мембрана выявляется только при электронной микроскопии. |
№ 11 Морфология грибов
Грибы относятся к царству Fungi (Mycetes, Mycota). Это многоклеточные или одноклеточные нефотосинтезирующие (бесхлорофильные) эукариотические микроорганизмы с клеточной стенкой. Грибы имеют ядро с ядерной оболочкой, цитоплазму с органеллами, цитоплазматическую мембрану и многослойную, ригидную клеточную стенку, состоящую из нескольких типов полисахаридов, а также белка, липидов и др. Некоторые грибы образуют капсулу. Цитоплазматическая мембрана содержит гликопротеины, фосфолипиды и эргостеролы. Грибы являются грамположительными микробами, вегетативные клетки — некислотоустойчивые. Грибы состоят из длинных тонких нитей (гиф), сплетающихся в грибницу, или мицелий. Гифы низших грибов — фикомицетов — не имеют перегородок. У высших грибов — эуми-цетов — гифы разделены перегородками; их мицелий многоклеточный. Различают гифальные и дрожжевые формы грибов. Гифальные (плесневые) грибы образуют ветвящиеся тонкие нити (гифы), сплетающиеся в грибницу, или мицелий (плесень). Гифы, врастающие в питательный субстрат, называются вегетативными гифами (отвечают за питание гриба), а растущие над поверхностью субстрата — воздушными или репродуктивными гифами (отвечают за бесполое размножение). Гифы низших грибов не имеют перегородок. Они представлены многоядерными клетками и называются ценоцитными. Гифы высших грибов разделены перегородками, или септами с отверстиями. Дрожжевые грибы (дрожжи), в основном, имеют вид отдельных овальных клеток (одноклеточные грибы). По типу полового размножения они распределены среди высших грибов — аскомицет и базидиомицет. При бесполом размножении дрожжи образуют почки или делятся, что приводит к одноклеточному росту. Могут образовывать псевдогифы и ложный мицелий (псевдомицелий) в виде цепочек удлиненных клеток — «сарделек». Грибы, аналогичные дрожжам, но не имеющие полового способа размножения, называют дрожжеподобными. Они размножаются только бесполым способом — почкованием или делением. Грибы размножаются спорами половым и бесполым способами, а также вегетативным путем (почкование или фрагментация гиф). Грибы, размножающиеся половым и бесполым путем, относятся к совершенным. Несовершенными называют грибы, у которых отсутствует или еще не описан половой путь размножения. Бесполое размножение осуществляется у грибов с помощью эндогенных спор, созревающих внутри круглой структуры — спорангия, и экзогенных спор — конидий, формирующихся на кончиках плодоносящих гиф. Типы грибов. Выделяют 3 типа грибов, имеющих половой способ размножения (так называемые совершенные грибы): зигомицеты (Zygomycota), аскомицеты (Ascomycota) и базидиомицеты (Basidiomycota). Отдельно выделяют условный, формальный тип/группу грибов — дейтеромицеты (Deiteromycota), у которых имеется только бесполый способ размножения (так называемые несовершенные грибы). |
|
|
|
|
|
№ 12 Морфология простейших
Простейшие — эукариотические одноклеточные микроорганизмы, составляющие подцарство Protozoa царства животных (Animalia). Простейшие включают 7 типов, из которых четыре типа (Sarcomastigophora, Apicomplexa, Ciliophora, Microspora) имеют представителей, вызывающих заболевания у человека. Размеры простейших колеблются в среднем от 5 до 30 мкм. Снаружи простейшие окружены мембраной (пелликулой) — аналогом цитоплазматической мембраны клеток животных. Некоторые простейшие имеют опорные фибриллы. Цитоплазма и ядро соответствуют по строению эукариотическим клеткам: цитоплазма состоит из эндоплазматического ретикулума, митохондрий, лизосом, многочисленных рибосом и др.; ядро имеет ядрышко и ядерную оболочку. Передвигаются простейшие посредством жгутиков, ресничек и путем образования псевдоподий. Простейшие могут питаться в результате фагоцитоза или образования особых структур. Многие простейшие при неблагоприятных условиях образуют цисты — покоящиеся стадии, устойчивые к изменению температуры, влажности и др. Простейшие окрашиваются по Романовскому—Гимзе (ядро — красного, цитоплазма — синего цвета). Тип Sarcomastigophora. Подтип Mastigophora (жгутиконосцы) включает следующих патогенных представителей: трипаносому — возбудителя африканского трипаносомоза (сонная болезнь); лейшмании — возбудителей кожной и висцеральной форм лейшманиозов; трихомонады, передающиеся половым путем и паразитирующие в толстой кишке человека; лямблию — возбудителя лямблиоза. Эти простейшие характеризуются наличием жгутиков: один — у лейшмании, четыре свободных жгутика и короткая ундулирующая мембрана — у три-хомонад. К подтипу Sarcodina (саркодовые) относится дизентерийная амеба — возбудитель амебной дизентерии человека. Морфологически сходна с ней непатогенная кишечная амеба. Эти простейшие передвигаются путем образования псевдоподий. Питательные вещества захватываются и погружаются в цитоплазму клеток. Половой путь размножения у амеб отсутствует. При неблагоприятных условиях они образуют цисту. Тип Apicomplexa. В классе Sporozoa (споровики) патогенными представителями являются возбудители токсоплазмоза, кокцидиоза, саркоцистоза и малярии. Жизненный цикл возбудителей малярии характеризуется чередованием полового размножения (в организме комаров Anopheles) и бесполого (в клетках тканей и эритроцитах человека они размножаются путем множественного деления). Токсоплазмы имеют форму полулуний. Токсоплазмозом человек заражается от животных. Токсоплазмы могут передаваться через плаценту и поражать центральную нервную систему и глаза плода. Тип Ciliophora. Патогенный представитель — возбудитель ба-лантидиаза — поражает толстый кишечник человека. Балантидии имеют многочисленные реснички и поэтому подвижны. Тип Microspora включает микроспоридии — маленькие (0,5—10 мкм) облигатные внутриклеточные паразиты, широко распространенные среди животных и вызывающие у ослабленных людей диарею и гнойно-воспалительные заболевания. |
№ 13 Особенности биологии вирусов
Вирусы — мельчайшие микробы, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только ДНК или РНК. Относятся к царству Vira. Являясь облигатными внутриклеточными паразитами, вирусы размножаются в цитоплазме или ядре клетки. Они — автономные генетические структуры. Отличаются особым — разобщенным (дисъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки, затем происходит их сборка в вирусные частицы. Сформированная вирусная частица называется вирионом. Морфологию и структуру вирусов изучают с помощью электронного микроскопа, так как их размеры малы и сравнимы с толщиной оболочки бактерий. Форма вирионов может быть различной: палочковидной (вирус табачной мозаики), пулевидной (вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), в виде сперматозоида (многие бактериофаги). Различают просто устроенные и сложно устроенные вирусы. Простые, или безоболочечные, вирусы состоят из нуклеиновой кислоты и белковой оболочки, называемой капсидом. Капсид состоит из повторяющихся морфологических субъединиц — капсомеров. Нуклеиновая кислота и капсид взаимодействуют друг с другом, образуя нуклеокапсид. Сложные, или оболочечные, вирусы снаружи капсида окружены ли-попротеиновой оболочкой (суперкапсидом, или пеплосом). Эта оболочка является производной структурой от мембран вирус-инфицированной клетки. На оболочке вируса расположены гликопротеиновые шипы, или шипики (пепломеры). Под оболочкой некоторых вирусов находится матриксный М-белок. Тип симметрии. Капсид или нуклеокапсид могут иметь спиральный, икосаэдрический (кубический) или сложный тип симметрии. Икосаэдрический тип симметрии обусловлен образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту (например, у вирусов гепатита А, герпеса, полиомиелита). Спиральный тип симметрии обусловлен винтообразной структурой нуклеокапсида (например, у вируса гриппа). Включения — скопление вирионов или отдельных их компонентов в цитоплазме или ядре клеток, выявляемые под микроскопом при специальном окрашивании. Вирус натуральной оспы образует цитоплазмати-ческие включения — тельца Гварниери; вирусы герпеса и аденовирусы — внутриядерные включения. Размеры вирусов определяют с помощью электронной микроскопии, методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Одним из самых мелких вирусов является вирус полиомиелита (около 20 нм), наиболее крупным — натуральной оспы (около 350 нм). Вирусы имеют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК-содержащие вирусы. Они обычно гаплоидны, т.е. имеют один набор генов. Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными. Среди РНК-содержащих вирусов различают вирусы с положительным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК). Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию. Вирусы поражают позвоночных и беспозвоночных животных, а также растения и бактерии. Являясь основными возбудителями инфекционных заболеваний человека, вирусы также участвуют в процессах канцерогенеза, могут передаваться различными путями, в том числе через плаценту (вирус краснухи, цитомега ловирус и др.), поражая плод человека. Они могут приводить к постинфекционным осложнениям — развитию миокардитов, панкреатитов, иммунодефицитов и др. Кроме обычных вирусов, известны и так называемые неканонические вирусы — прионы — белковые инфекционные частицы, являющиеся агентами белковой природы, имеющие вид фибрилл размером 10—20x100—200 нм. Прионы, по-видимому, являются одновременно индукторами и продуктами автономного гена человека или животного и вызывают у них энцефалопатии в условиях медленной вирусной инфекции (болезни Крейтц-фельдта—Якоба, куру и др.). Другими необычными агентами, близкими к вирусам, являются вироиды — небольшие молекулы кольцевой, суперспи-рализованной РНК, не содержащие белка, вызывающие заболевания у растений. |
№ 14 Структура и химический состав вирусов и бактериофагов
Вирусы — мельчайшие микробы, не имеющие клеточного строения, белоксинтезирующей системы, содержащие только ДНК или РНК. Относятся к царству Vira. Являясь облигатными внутриклеточными паразитами, вирусы размножаются в цитоплазме или ядре клетки. Они — автономные генетические структуры. Отличаются особым — разобщенным (дисъюнктивным) способом размножения (репродукции): в клетке отдельно синтезируются нуклеиновые кислоты вирусов и их белки, затем происходит их сборка в вирусные частицы. Сформированная вирусная частица называется вирионом. Морфологию и структуру вирусов изучают с помощью электронного микроскопа, так как их размеры малы и сравнимы с толщиной оболочки бактерий. Форма вирионов может быть различной: палочковидной (вирус табачной мозаики), пулевидной (вирус бешенства), сферической (вирусы полиомиелита, ВИЧ), в виде сперматозоида (многие бактериофаги). Различают просто устроенные и сложно устроенные вирусы. Простые, или безоболочечные, вирусы состоят из нуклеиновой кислоты и белковой оболочки, называемой капсидом. Капсид состоит из повторяющихся морфологических субъединиц — капсомеров. Нуклеиновая кислота и капсид взаимодействуют друг с другом, образуя нуклеокапсид. Сложные, или оболочечные, вирусы снаружи капсида окружены ли-попротеиновой оболочкой (суперкапсидом, или пеплосом). Эта оболочка является производной структурой от мембран вирус-инфицированной клетки. На оболочке вируса расположены гликопротеиновые шипы, или шипики (пепломеры). Под оболочкой некоторых вирусов находится матриксный М-белок. Капсид и суперкапсид защищают вирионы от влияния окружающей среды, обусловливают избирательное взаимодействие (адсорбцию) с клетками, определяют антигенные и иммуногенные свойства вирионов. Внутренние структуры вирусов называются сердцевиной. Тип симметрии. Капсид или нуклеокапсид могут иметь спиральный, икосаэдрический (кубический) или сложный тип симметрии. Икосаэдрический тип симметрии обусловлен образованием изометрически полого тела из капсида, содержащего вирусную нуклеиновую кислоту (например, у вирусов гепатита А, герпеса, полиомиелита). Спиральный тип симметрии обусловлен винтообразной структурой нуклеокапсида (например, у вируса гриппа). Включения — скопление вирионов или отдельных их компонентов в цитоплазме или ядре клеток, выявляемые под микроскопом при специальном окрашивании. Вирус натуральной оспы образует цитоплазмати-ческие включения — тельца Гварниери; вирусы герпеса и аденовирусы — внутриядерные включения. Размеры вирусов определяют с помощью электронной микроскопии, методом ультрафильтрации через фильтры с известным диаметром пор, методом ультрацентрифугирования. Одним из самых мелких вирусов является вирус полиомиелита (около 20 нм), наиболее крупным — натуральной оспы (около 350 нм). Вирусы имеют уникальный геном, так как содержат либо ДНК, либо РНК. Поэтому различают ДНК-содержащие и РНК-содержащие вирусы. Они обычно гаплоидны, т.е. имеют один набор генов. Геном вирусов представлен различными видами нуклеиновых кислот: двунитчатыми, однонитчатыми, линейными, кольцевыми, фрагментированными. Среди РНК-содержащих вирусов различают вирусы с положительным (плюс-нить РНК) геномом. Плюс-нить РНК этих вирусов выполняет наследственную функцию и функцию информационной РНК (иРНК). Имеются также РНК-содержащие вирусы с отрицательным (минус-нить РНК) геномом. Минус-нить РНК этих вирусов выполняет только наследственную функцию. Геном вирусов способен включаться в состав генетического аппарата клетки в виде провируса, проявляя себя генетическим паразитом клетки. Нуклеиновые кислоты некоторых вирусов (вирусы герпеса и др.) могут находиться в цитоплазме инфицированных клеток, напоминая плазмиды. |
|
|
||
|
|
|
№ 15 Методы микроскопии (люминесцентная, темнопольная, фазово-контрастная, электронная).
Люминесцентная (или флюоресцентная) микроскопия. Основана на явлении фотолюминесценции. Люминесценция — свечение веществ, возникающее после воздействия на них каких-либо источников энергии: световых, электронных лучей, ионизирующего излучения. Фотолюминесценция — люминесценция объекта под влиянием света. Если освещать люминесцирующий объект синим светом, то он испускает лучи красного, оранжевого, желтого или зеленого цвета. В результате возникает цветное изображение объекта. Темнопольная микроскопия. Микроскопия в темном поле зрения основана на явлении дифракции света при сильном боковом освещении взвешенных в жидкости мельчайших частиц (эффект Тиндаля). Эффект достигается с помощью параболоид- или кардиоидконденсора, которые заменяют обычный конденсор в биологическом микроскопе . Фазово-контрастная микроскопия. Фазово-контрастное приспособление дает возможность увидеть в микроскоп прозрачные объекты. Они приобретают высокую контрастность изображения, которая может быть позитивной или негативной. Позитивным фазовым контрастом называют темное изображение объекта в светлом поле зрения, негативным — светлое изображение объекта на темном фоне. Для фазово-контрастной микроскопии используют обычный микроскоп и дополнительное фазово-контрастное устройство, а также специальные осветители. Электронная микроскопия. Позволяет наблюдать объекты, размеры которых лежат за пределами разрешающей способности светового микроскопа (0,2 мкм). Электронный микроскоп применяется для изучения вирусов, тонкого строения различных микроорганизмов, макромолекулярных структур и других субмикроскопических объектов. |
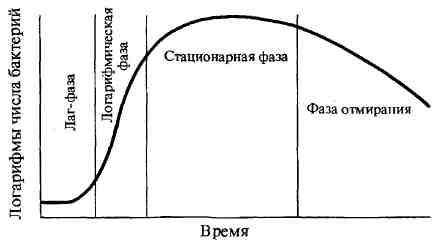
№ 16 Рост и размножение бактерий. Фазы размножения.
Жизнедеятельность бактерий характеризуется ростом — формированием структурно-функциональных компонентов клетки и увеличением самой бактериальной клетки, а также размножением — самовоспроизведением, приводящим к увеличению количества бактериальных клеток в популяции. Бактерии размножаются путем бинарного деления пополам, реже путем почкования. Актиномицеты, как и грибы, могут размножаться спорами. Актиномицеты, являясь ветвящимися бактериями, размножаются путем фрагментации нитевидных клеток. Грамположительные бактерии делятся путем врастания синтезирующихся перегородок деления внутрь клетки, а грамотрицательные — путем перетяжки, в результате образования гантелевид-ных фигур, из которых образуются две одинаковые клетки. Делению клеток предшествует репликация бактериальной хромосомы по полуконсервативному типу (двуспиральная цепь ДНК раскрывается и каждая нить достраивается комплементарной нитью), приводящая к удвоению молекул ДНК бактериального ядра — нуклеоида. Репликация ДНК происходит в три этапа: инициация, элонгация, или рост цепи, и терминация. Размножение бактерий в жидкой питательной среде. Бактерии, засеянные в определенный, не изменяющийся объем питательной среды, размножаясь, потребляют питательные элементы, что приводит в дальнейшем к истощению питательной среды и прекращению роста бактерий. Культивирование бактерий в такой системе называют периодическим культивированием, а культуру — периодической. Если же условия культивирования поддерживаются путем непрерывной подачи свежей питательной среды и оттока такого же объема культуральной жидкости, то такое культивирование называется непрерывным, а культура — непрерывной. При выращивании бактерий на жидкой питательной среде наблюдается придонный, диффузный или поверхностный (в виде пленки) рост культуры. Рост периодической культуры бактерий, выращиваемых на жидкой питательной среде, подразделяют на несколько фаз, или периодов: 1. лаг-фаза; 2. фаза логарифмического роста; 3. фаза стационарного роста, или максимальной концентрации бактерий; 4. фаза гибели бактерий. Эти фазы можно изобразить графически в виде отрезков кривой размножения бактерий, отражающей зависимость логарифма числа живых клеток от времени их культивирования.
Лаг-фаза — период между посевом бактерий и началом размножения. Продолжительность лаг-фазы в среднем 4—5 ч. Бактерии при этом увеличиваются в размерах и готовятся к делению; нарастает количество нуклеиновых кислот, белка и других компонентов. Фаза логарифмического (экспоненциального) роста является периодом интенсивного деления бактерий. Продолжительность ее около 5— 6 ч. При оптимальных условиях роста бактерии могут делиться каждые 20—40 мин. Во время этой фазы бактерии наиболее ранимы, что объясняется высокой чувствительностью компонентов метаболизма интенсивно растущей клетки к ингибиторам синтеза белка, нуклеиновых кислот и др. Затем наступает фаза стационарного роста, при которой количество жизнеспособных клеток остается без изменений, составляя максимальный уровень (М-концентрация). Ее продолжительность выражается в часах и колеблется в зависимости от вида бактерий, их особенностей и культивирования. Завершает процесс роста бактерий фаза гибели, характеризующаяся отмиранием бактерий в условиях истощения источников питательной среды и накопления в ней продуктов метаболизма бактерий. Продолжительность ее колеблется от 10 ч до нескольких недель. Интенсивность роста и размножения бактерий зависит от многих факторов, в том числе оптимального состава питательной среды, окислительно-восстановительного потенциала, рН, температуры и др. Размножение бактерий на плотной питательной среде. Бактерии, растущие на плотных питательных средах, образуют изолированные колонии округлой формы с ровными или неровными краями (S- и R-формы), различной консистенции и цвета, зависящего от пигмента бактерий. Пигменты, растворимые в воде, диффундируют в питательную среду и окрашивают её. Другая группа пигментов нерастворима в воде, но растворима в органических растворителях. И, наконец, существуют пигменты, не растворимые ни в воде, ни в органических соединениях. Наиболее распространены среди микроорганизмов такие пигменты, как каротины, ксантофиллы и меланины. Меланины являются нерастворимыми пигментами черного, коричневого или красного цвета, синтезирующимися из фенольных соединений. Меланины наряду с каталазой, супероксидцисмутазой и пероксидазами защищают микроорганизмы от воздействия токсичных перекисных радикалов кислорода. Многие пигменты обладают антимикробным, антибиотикоподобным действием. |
№ 17 Способы получения энергии бактериями (дыхание, брожение). Методы культивирования анаэробов.
Дыхание, или биологическое окисление, основано на окислительно-восстановительных реакциях, идущих с образованием АТФ-универсального аккумулятора химической энергии. Энергия необходима микробной клетке для ее жизнедеятельности. При дыхании происходят процессы окисления и восстановления: окисление — отдача донорами (молекулами или атомами) водорода или электронов; восстановление — присоединение водорода или электронов к акцептору. Акцептором водорода или электронов может быть молекулярный кислород (такое дыхание называется аэробным) или нитрат, сульфат, фумарат (такое дыхание называется анаэробным — нитратным, сульфатным, фумаратным). Анаэробиоз (от греч. аег — воздух + bios — жизнь) — жизнедеятельность, протекающая при отсутствии свободного кислорода. Если донорами и акцепторами водорода являются органические соединения, то такой процесс называется брожением. При брожении происходит ферментативное расщепление органических соединений, преимущественно углеводов, в анаэробных условиях. С учетом конечного продукта расщепления углеводов различают спиртовое, молочнокислое, уксуснокислое и другие виды брожения. По отношению к молекулярному кислороду бактерии можно разделить на три основные группы: облигатные, т.е. обязательные, аэробы, облигатные анаэробы и факультативные анаэробы. Методы культивирования анаэробов. Для культивирования анаэробов необходимо понизить окислительно-восстановительный потенциал среды, создать условия анаэробиоза, т. е. пониженного содержания кислорода в среде и окружающем ее пространстве. Это достигается применением физических, химических и биологических методов. Физические методы. Основаны на выращивании микроорганизмов в безвоздушной среде, что достигается: 1) посевом в среды, содержащие редуцирующие и легко окисляемые вещества; 2) посевом микроорганизмов в глубину плотных питательных сред; 3) механическим удалением воздуха из сосудов, в которых выращиваются анаэробные микроорганизмы; 4) заменой воздуха в сосудах каким-либо индифферентным газом. В качестве редуцирующих веществ обычно используют кусочки (около 0,5 г) животных или растительных тканей (печень, мозг, почки, селезенка, кровь, картофель, вата). Эти ткани связывают растворенный в среде кислород и адсорбируют бактерии. Чтобы уменьшить содержание кислорода в питательной среде, ее перед посевом кипятят 10—15 мин, а затем быстро охлаждают и заливают сверху небольшим количеством стерильного вазелинового масла. Высота слоя масла в пробирке около 1 см. В качестве легко окисляемых веществ используют глюкозу, лактозу и муравьинокислый натрий. Лучшей жидкой питательной средой с редуцирующими веществами является среда Китта — Тароцци, которая используется с успехом для накопления анаэробов при первичном посеве из исследуемого материала и для поддержания роста выделенной чистой культуры анаэробов. Посев микроорганизмов в глубину плотных сред производят по способу Виньяль — Вейона, который состоит в механической защите посевов анаэробов от кислорода воздуха. Берут стеклянную трубку длиной 30 см и диаметром 3—6 мм. Один конец трубки вытягивают в капилляр в виде пастеровской пипетки, а у другого конца делают перетяжку. В оставшийся широкий конец трубки вставляют ватную пробку. В пробирки с расплавленным и охлажденным до 50°С питательным агаром засевают исследуемый материал. Затем насасывают засеянный агар в стерильные трубки Виньяль — Вейона. Капиллярный конец трубки запаивают в пламени горелки и трубки помещают в термостат. Так создаются благоприятные условия для роста самых строгих анаэробов. Для выделения отдельной колонии трубку надрезают напильником, соблюдая правила асептики, на уровне колонии, ломают, а колонию захватывают стерильной петлей и переносят в пробирку с питательной средой для дальнейшего выращивания и изучения в чистом виде. Удаление воздуха производят путем его механического откачивания из специальных приборов — анаэроста-тов, в которые помещают чашки с посевом анаэробов. Переносный анаэростат представляет собой толстостенный металлический цилиндр с хорошо притертой крышкой (с резиновой прокладкой), снабженный отводящим краном и вакуумметром. После размещения засеянных чашек или пробирок воздух из анаэростата удаляют с помощью вакуумного насоса. Замену воздуха индифферентным газом (азотом, водородом, аргоном, углекислым газом) можно производить в тех же анаэростатах путем вытеснения его газом из баллона. Химические методы. Основаны на поглощении кислорода воздуха в герметически закрытом сосуде (анаэро-стате, эксикаторе) такими веществами, как пирогаллол или гидросульфит натрия Na2S204. Биологические методы. Основаны на совместном выращивании анаэробов со строгими аэробами. Для этого из застывшей агаровой пластинки по диаметру чашки вырезают стерильным скальпелем полоску агара шириной около 1 см. Получается два агаровых полудиска в одной чашке. На одну сторону агаровой пластинки засевают аэроб, например часто используют S. aureus или Serratia marcescens. На другую сторону засевают анаэроб. Края чашки заклеивают пластилином или заливают расплавленным парафином и помещают в термостат. При наличии подходящих условий в чашке начнут размножаться аэробы. После того, как весь кислород в пространстве чашки будет ими использован, начнется рост анаэробов (через 3—4 сут). В целях сокращения воздушного пространства в чашке питательную среду наливают возможно более толстым слоем. Комбинированные методы. Основаны на сочетании физических, химических и биологических методов создания анаэробиоза. |
|
№ 18 Типы и механизмы питания бактерий.
Типы питания. Микроорганизмы нуждаются в углеводе, азоте, сере, фосфоре, калии и других элементах. В зависимости от источников углерода для питания бактерии делятся на аутотрофы, использующие для построения своих клеток диоксид углерода С02 и другие неорганические соединения, и гетеротрофы, питающиеся за счет готовых органических соединений. Аутотрофными бактериями являются нитрифицирующие бактерии, находящиеся в почве; серобактерии, обитающие в воде с сероводородом; железобактерии, живущие в воде с закисным железом, и др. Гетеротрофы, утилизирующие органические остатки отмерших организмов в окружающей среде, называются сапрофитами. Гетеротрофы, вызывающие заболевания у человека или животных, относят к патогенным и условно-патогенным. Среди патогенных микроорганизмов встречаются облигатные и факультативные паразиты (от греч. parasitos — нахлебник). Облигатные паразиты способны существовать только внутри клетки, например риккетсии, вирусы и некоторые простейшие. В зависимости от окисляемого субстрата, называемого донором электронов или водорода, микроорганизмы делят на две группы. Микроорганизмы, использующие в качестве доноров водорода неорганические соединения, называют литотрофны-ми (от греч. lithos — камень), а микроорганизмы, использующие в качестве доноров водорода органические соединения, — органотрофами. Учитывая источник энергии, среди бактерий различают фототрофы, т.е. фотосинтезирующие (например, сине-зеленые водоросли, использующие энергию света), и хемотрофы, нуждающиеся в химических источниках энергии. Механизмы питания. Поступление различных веществ в бактериальную клетку зависит от величины и растворимости их молекул в липидах или воде, рН среды, концентрации веществ, различных факторов проницаемости мембран и др. Клеточная стенка пропускает небольшие молекулы и ионы, задерживая макромолекулы массой более 600 Д. Основным регулятором поступления веществ в клетку является цитоплазматическая мембрана. Условно можно выделить четыре механизма проникновения питательных веществ в бактериальную клетку: это простая диффузия, облегченная диффузия, активный транспорт, транслокация групп. Наиболее простой механизм поступления веществ в клетку — простая диффузия, при которой перемещение веществ происходит вследствие разницы их концентрации по обе стороны цитоплазматической мембраны. Вещества проходят через липид-ную часть цитоплазматической мембраны (органические молекулы, лекарственные препараты) и реже по заполненным водой каналам в цитоплазматической мембране. Пассивная диффузия осуществляется без затраты энергии. Облегченная диффузия происходит также в результате разницы концентрации веществ по обе стороны цитоплазматической мембраны. Однако этот процесс осуществляется с помощью молекул-переносчиков, локализующихся в цитоплазматической мембране и обладающих специфичностью. Каждый переносчик транспортирует через мембрану соответствующее вещество или передает другому компоненту цитоплазматической мембраны — собственно переносчику. Белками-переносчиками могут быть пермеазы, место синтеза которых — цитоплазматическая мембрана. Облегченная диффузия протекает без затраты энергии, вещества перемещаются от более высокой концентрации к более низкой. Активный транспорт происходит с помощью пермеаз и направлен на перенос веществ от меньшей концентрации в сторону большей, т.е. как бы против течения, поэтому данный про цесс сопровождается затратой метаболической энергии (АТФ), образующейся в результате окислительно-восстановительных реакций в клетке. Перенос (транслокация) групп сходен с активным транспортом, отличаясь тем, что переносимая молекула видоизменяется в процессе переноса, например фосфорилируется. Выход веществ из клетки осуществляется за счет диффузии и при участии транспортных систем. |
||
|
|
|
|
|
№ 19 Основные принципы культивирования бактерий.
Универсальным инструментом для производства посевов является бактериальная петля. Кроме нее, для посева уколом применяют специальную бактериальную иглу, а для посевов на чашках Петри — металлические или стеклянные шпатели. Для посевов жидких материалов наряду с петлей используют пастеровские и градуированные пипетки. Первые предварительно изготовляют из стерильных легкоплавких стеклянных трубочек, которые вытягивают на пламени в виде капилляров. Конец капилляра сразу же запаивают для сохранения стерильности. У пастеровских и градуированных пипеток широкий конец закрывают ватой, после чего их помещают в специальные пеналы или обертывают бумагой и стерилизуют. При пересеве бактериальной культуры берут пробирку в левую руку, а правой, обхватив ватную пробку IV и V пальцами, вынимают ее, пронося над пламенем горелки. Удерживая другими пальцами той же руки петлю, набирают ею посевной материал, после чего закрывают пробирку пробкой. Затем в пробирку со скошенным агаром вносят петлю с посевным материалом, опуская ее до конденсата в нижней части среды, и зигзагообразным движением распределяют мате риал по скошенной поверхности агара. Вынув петлю, обжигают край пробирки и закрывают ее пробкой. Петлю стерилизуют в пламени горелки и ставят в штатив. Пробирки с посевами надг писывают, указывая дату посева и характер посевного материала (номер исследования или название культуры). Посевы «газоном» производят шпателем на питательный агар в чашке Петри. Для этого, приоткрыв левой рукой крышку, петлей или пипеткой наносят посевной материал на поверхность питательного агара. Затем проводят шпатель через пламя горелки, остужают его о внутреннюю сторону крышки и растирают материал по всей поверхности среды. После инкубации посева появляется равномерный сплошной рост бактерий. |
|
|
|
№ 20 Искусственные питательные среды, их классификация. Требования, предъявляемые к питательным средам.
Питательной средой в микробиологии называют среды, содержащие различные соединения сложного или простого состава, которые применяются для размножения бактерий или других микроорганизмов в лабораторных или промышленных условиях. Питательные среды готовят из продуктов животного или растительного происхождения. Большое значение имеет наличие в питательной среде ростовых факторов, которые катализируют метаболические процессы микробной клетки (витамины группы В, никотиновая кислота и др.). Искусственные среды готовят по определенным рецептам из различных настоев или отваров животного или растительного происхождения с добавлением неорганических солей, углеводов и азотистых веществ. В бактериологической практике чаще всего используют сухие питательные среды, которые получают на основе достижений современной биотехнологии. Для их приготовления используют экономически рентабельное непищевое сырье: утратившие срок годности кровезаменители (гидролизин—кислотный гидролизат крови животных, аминопептид — ферментативный гидролизат крови; продукты биотехнологии (кормовые дрожжи, кормовой лизин, виноградная мука, белколизин). Сухие питательные среды могут храниться в течение длительного времени, удобны при транспортировке и имеют относительно стандартный состав. По консистенции питательные среды могут быть жидкими, полужидкими, плотными. Плотные среды готовят путем добавления к жидкой среде 1,5—2% агара, полужидкие — 0,3— 0,7 % агара. Агар представляет собой продукт переработки особого вида морских водорослей, он плавится при температуре 80—86 °С, затвердевает при температуре около 40 °С и в застывшем состоянии придает среде плотность. В некоторых случаях для получения плотных питательных сред используют желатин (10—15%). Ряд естественных питательных сред (свернутая сыворотка крови, свернутый яичный белок) сами по себе являются плотными. По целевому назначению среды подразделяют на основные, элективные и дифференциально-диагностические. К основным относятся среды, применяемые для выращивания многих бактерий. Это триптические гидролизаты мясных, рыбных продуктов, крови животных или казеина, из которых готовят жидкую среду — питательный бульон и плотную — питательный агар. Такие среды служат основой для приготовления сложных питательных сред — сахарных, кровяных и др., удовлетворяющих пищевые потребности патогенных бактерий. Элективные питательные среды предназначены для избирательного выделения и накопления микроорганизмов определенного вида (или определенной группы) из материалов, содержащих разнообразную постороннюю микрофлору. При создании элективных питательных сред исходят из биологических особенностей, которые отличают данные микроорганизмы от большинства других. Например, избирательный рост стафилококков наблюдается при повышенной концентрации хлорида натрия, холерного вибриона — в щелочной среде и т. д. Дифференциально-диагностические питательные среды применяются для разграничения отдельных видов (или групп) микроорганизмов. Принцип построения этих сред основан на том, что разные виды бактерий различаются между собой по биохимической активности вследствие неодинакового набора ферментов. Особую группу составляют синтетические и полусинтетические питательные среды. В состав синтетических сред входят химически чистые вещества: аминокислоты, минеральные соли, углеводы, витамины. В полусинтетические среды дополнительно включают пептон, дрожжевой экстракт и другие питательные вещества. Эти среды чаще всего применяют в научно-исследовательской работе и в микробиологической промышленности при получении антибиотиков, вакцин и других препаратов. В последние годы в целях экономии питательных сред и ускоренной идентификации некоторых микроорганизмов (энтеробактерии, стафилококки, стрептококки и др.) применяются так называемые микротест-системы (МТС). Они представляют собой полистироловые пластины с лунками, в которых содержатся стерильные дифференциально-диагностические среды. Стерилизацию МТС проводят УФ-облучением. Микротест-системы особенно удобны при массовых бактериологических исследованиях в практических лабораториях. Требования, предъявляемые к питательным средам. Любая питательная среда должна отвечать следующим требованиям: содержать все необходимые для размножения микроорганизмов вещества в легкоусвояемой форме; иметь оптимальные влажность, вязкость, рН, быть изотоничной и по возможности прозрачной. Каждую питательную среду стерилизуют определенным способом в зависимости от ее состава. |
№ 21 Принципы и методы выделения чистых культур бактерий.
Чистой культурой называется популяция бактерий одного вида или одной разновидности, выращенная на питательной среде. Многие виды бактерий подразделяют по одному признаку на биологические варианты — биовары. Биовары, различающиеся по биохимическим свойствам, называют хемоварами, по антигенным свойствам — сероварами, по чувствительности к фагу — фаговарами. Культуры микроорганизмов одного и того же вида, или биовара, выделенные из различных источников или в разное время из одного и того же источника, называют штаммами, которые обычно обозначаются номерами или какими-либо символами. Чистые культуры бактерий в диагностических бактериологических лабораториях получают из изолированных колоний, пересевая их петлей в пробирки с твердыми или, реже, жидкими питательными средами. Колония представляет собой видимое изолированное скопление особей одного вида микроорганизмов, образующееся в результате размножения одной бактериальной клетки на плотной питательной среде (на поверхности или в глубине ее). Колонии бактерий разных видов отличаются друг от друга по своей морфологии, цвету и другим признакам. Чистую культуру бактерий получают для проведения диагностических исследований — идентификации, которая достигается путем определения морфологических, культуральных, биохимических и других признаков микроорганизма. Морфологические и тинкториальные признаки бактерий изучают при микроскопическом исследовании мазков, окрашенных разными методами, и нативных препаратов. Культуральные свойства характеризуются питательными потребностями, условиями и типом роста бактерий на плотных и жидких питательных средах. Они устанавливаются по морфологии колоний и особенностям роста культуры. Биохимические признаки бактерий определяются набором конститутивных и индуцибельных ферментов, присущих определенному роду, виду, варианту. В бактериологической практике таксономическое значение имеют чаще всего сахаролитические и протеолитические ферменты бактерий, которые определяют на дифференциально-диагностических средах. При идентификации бактерий до рода и вида обращают внимание на пигменты, окрашивающие колонии и культуральную среду в разнообразные цвета. Например, красный пигмент образуют Serratia marcescens, золотистый пигмент — Staphylococcus aureus (золотистый стафилококк), сине-зеленый пигмент — Pseu-domonas aeruginosa. Для установления биовара (хемовара, серовара, фаготипа) проводят дополнительные исследования по выялвениб соответствующего маркера – определению фермента, антигена, чувствительности к Фанам. Методы выделения чистых культур бактерий. Универсальным инструментом для производства посевов является бактериальная петля. Кроме нее, для посева уколом применяют специальную бактериальную иглу, а для посевов на чашках Петри — металлические или стеклянные шпатели. Для посевов жидких материалов наряду с петлей используют пастеровские и градуированные пипетки. Первые предварительно изготовляют из стерильных легкоплавких стеклянных трубочек, которые вытягивают на пламени в виде капилляров. Конец капилляра сразу же запаивают для сохранения стерильности. У пастеровских и градуированных пипеток широкий конец закрывают ватой, после чего их помещают в специальные пеналы или обертывают бумагой и стерилизуют. При пересеве бактериальной культуры берут пробирку в левую руку, а правой, обхватив ватную пробку IV и V пальцами, вынимают ее, пронося над пламенем горелки. Удерживая другими пальцами той же руки петлю, набирают ею посевной материал, после чего закрывают пробирку пробкой. Затем в пробирку со скошенным агаром вносят петлю с посевным материалом, опуская ее до конденсата в нижней части среды, и зигзагообразным движением распределяют мате риал по скошенной поверхности агара. Вынув петлю, обжигают край пробирки и закрывают ее пробкой. Петлю стерилизуют в пламени горелки и ставят в штатив. Пробирки с посевами надг писывают, указывая дату посева и характер посевного материала (номер исследования или название культуры). Посевы «газоном» производят шпателем на питательный агар в чашке Петри. Для этого, приоткрыв левой рукой крышку, петлей или пипеткой наносят посевной материал на поверхность питательного агара. Затем проводят шпатель через пламя горелки, остужают его о внутреннюю сторону крышки и растирают материал по всей поверхности среды. После инкубации посева появляется равномерный сплошной рост бактерий. |
№ 22 Ферменты бактерий. Идентификация бактерий по ферментативной активности.
В основе всех метаболических реакций в бактериальной клетке лежит деятельность ферментов, которые принадлежат к 6 классам: оксиредуктазы, трансферазы, гидролазы, лигазы, лиазы, изомеразы. Ферменты, образуемые бактериальной клеткой, могут локализоваться как внутри клетки — эндоферменты, так и выделяться в окружающую среду — экзоферменты. Экзоферменты играют большую роль в обеспечении бактериальной клетки доступными для проникновения внутрь источниками углерода и энергии. Большинство гидролаз является экзоферментами, которые, выделяясь в окружающую среду, расщепляют крупные молекулы пептидов, полисахаридов, липидов до мономеров и димеров, способных проникнуть внутрь клетки. Ряд экзоферментов, например гиалуронидаза, коллагеназа и другие, являются ферментами агрессии. Некоторые ферменты локализованы в периплазматическом пространстве бактериальной клетки. Они участвуют в процессах переноса веществ в бактериальную клетку. Ферментативный спектр является таксономическим признаком, характерным для семейства, рода и — в некоторых случаях — для видов. Поэтому определением спектра ферментативной активности пользуются при установлении таксономического положения бактерий. Наличие экзоферментов можно определить при помощи дифференциально-диагностических сред, поэтому для идентификации бактерий разработаны специальные тест-системы, состоящие из набора дифференциально-диагностических сред. Идентификация бактерий по ферментативной активности. Наиболее часто определяют ферменты класса гидролаз и оксидоредуктаз, используя специальные методы и среды. Для определения протеолитической активности микроорганизмы засевают в столбик желатина уколом. Через 3—5 дней посевы просматривают и отмечают характер разжижения желатина. При разложении белка некоторыми бактериями могут выделяться специфические продукты — индол, сероводород, аммиак. Для их определения служат специальные индикаторные бумажки, которые помещают между горлышком и ватной пробкой в пробирку с МПБ или (и) пептонной водой, засеянными изучаемыми микроорганизмами. Индол (продукт разложения триптофана) окрашивает в розовый цвет полоску бумаги, пропитанной насыщенным раствором щавелевой кислоты. Бумага, пропитанная раствором ацетата свинца, в присутствии сероводорода чернеет. Для определения аммиака используют красную лакмусовую бумажку. Для многих микроорганизмов таксономическим признаком служит способность разлагать определенные углеводы с образованием кислот и газообразных продуктов. Для выявления этого используют среды Гисса, содержащие различные углеводы (глюкозу, сахарозу, мальтозу, лактозу и др.). Для обнаружения кислот в среду добавлен реактив Андреде, который изменяет свой цвет от бледно-желтого до красного в интервале рН 7,2—6,5, поэтому набор сред Гисса с ростом микроорганизмов называют «пестрым рядом». Для обнаружения газообразования в жидкие среды опускают поплавки или используют полужидкие среды с 0,5% агара. Для того чтобы определить интенсивное кислотообразование, характерное для брожения смешанного типа, в среду с 1% глюкозы и 0,5% пептона (среда Кларка) добавляют индикатор метиловый красный, который имеет желтый цвет при рН 4,5 и выше, и красный —при более низких значениях рН. Гидролиз мочевины определяют по выделению аммиака (лакмусовая бумажка) и подщелачиванию среды. При идентификации многих микроорганизмов используют реакцию Фогеса — Проскауэра на ацетоин — промежуточное соединение при образовании бутандиола из пировиноградной кислоты. Положительная реакция свидетельствует о наличии бутандиолового брожения. Обнаружить каталазу можно по пузырькам кислорода, которые начинают выделяться сразу же после смешивания микробных клеток с 1 % раствором перекиси водорода. Для определения цитохромоксидазы применяют реактивы: 1) 1% спиртовый раствор сс-нафтола-1; 2) 1% водный раствор N-диметил-р-фенилендиамина дигидро-хлорида. О наличии цитохромоксидазы судят по синему окрашиванию, появляющемуся через 2—5 мин. Для определения нитритов используют реактив Грисса: Появление красного окрашивания свидетельствует о наличии нитритов. |
|
|
||
|
|
||
|
№ 23 Внутривидовая идентификация бактерий (эпидемиологическое маркирование).
С целью выявления эпидемической цепочки заболевания, в т. ч. для обнаружения источника инфекции, осуществляют внутривидовую идентификацию бактерий, к-рая заключается в определении фаготипа (фаговара), изучении антигенных и других свойств выделенных бактерий. Определение фаготипа - фаготипирование производят при стафилококковой инфекции, брюшном тифе, паратифе В. Фаготипирование — один из методов эпидемиологического маркирования. Применяется для выявления источника инфекции. Выделение бактерий одного фаговара от разных больных указывает на общий источник их заражения. Предварительно фаготируется. При внутривидовой идентификации бактерий, т. е. при определении фаговара (фаготипа) бактерий с помощью фаготи-пирования, на чашку с плотной питательной средой, засеянную чистой культурой возбудителя в виде «газона», наносят капли различных диагностических типоспецифических фагов. Бактерии, чувствительные к фагу, лизируются (образуется стерильное пятно, «бляшка» или так называемая негативная колония фага). На засеянные «газоном» стафилококки наносятся капли взвеси стафилококковых бактериофагов. Через сутки после инкубации в термостате видны стерильные зоны отсутствия роста бактерий (стерильные «бляшки») в результате размножения бактериофагов, вызывающих лизис этих бактерий. |
№ 24 Особенности физиологии грибов.
Грибы относятся к царству Fungi (Mycetes, Mycota). Это многоклеточные или одноклеточные нефотосинтезирующие (бес-хлорофильные) эукариотические микроорганизмы с клеточной стенкой. Грибы по типу питания — гетеротрофы, по отношению к кислороду — аэробы и факультативные анаэробы. Растут в широких диапазонах температур (оптимальная температура 25—30 °С), имеют половой и бесполый способы размножения. Поэтому грибы широко распространены в окружающей среде, особенно в почве. Грибы вместе с сине-зелеными водорослями образуют симбиоз в виде лишайника. В этом симбиозе грибы поглощают воду и растворимые в ней вещества, а сине-зеленые водоросли поставляют грибам органические соединения. Другой вид взаимоотношений — микориза — симбиоз грибов и корней высших растений. Грибы культивируют в течение нескольких суток на сусле-агаре или жидком сусле, среде Сабуро, Чапека и др. Для этой цели можно использовать лабораторных животных. Некоторые грибы обладают диморфизмом, т. е. способностью образовывать нитчатые и дрожжевые формы в зависимости от условий роста. Дрожжеподобные формы часто образуются in vivo, т. е. при инфицировании человека грибами. Размножение грибов происходит половым и бесполым (вегетативным) способами. Половое размножение грибов происходит с образованием гамет, половых спор и других половых форм. Половые формы называются телеоморфами. Бесполое (вегетативное) размножение грибов происходит с образованием соответствующих форм, называемых анаморфами. Типы грибов. Выделяют 3 типа грибов, имеющих половой способ размножения (так называемые совершенные грибы): зигомицеты (Zygomycota), аскомицеты (Ascomycota) и базидиомицеты (Basidiomycota). Отдельно выделяют условный, формальный тип/группу грибов — дейтеромицеты (Deiteromycota), у которых имеется только бесполый способ размножения (так называемые несовершенные грибы). |
№ 25 Особенности физиологии простейших.
Простейшие — эукариотические одноклеточные микроорганизмы, составляющие подцарство Protozoa в царстве животных (Animalia); являются одноклеточными животными. Снаружи простейшие окружены мембраной (пелликулой) — аналогом цитоплазматической мембраны клеток животных. Они содержат: ядро с ядерной оболочкой и ядрышком; цитоплазму, состоящую из эндоплазматического ретикулума, митохондрий, лизосом, многочисленных рибосом и др. Размеры простейших колеблются в среднем от 2 до 100 мкм. Снаружи они окружены мембраной (пелликулой) — аналогом цитоплазматической мембраны клеток животных. Простейшие представлены 7 типами, из которых четыре типа (Sarcomastigophora, Apicomplexa, Ciliopkora, Microspora) включают возбудителей заболеваний у человека. Простейшие имеют: органы движения (жгутики, реснички, псевдоподии), питания (пищеварительные вакуоли) и выделения (сократительные вакуоли); могут питаться в результате фагоцитоза или образования особых структур. Некоторые простейшие имеют опорные фибриллы. Размножаются бесполым путем — двойным делением или множественным делением (шизогония), а некоторые и половым путем (спорогония). Многие из них при неблагоприятных условиях образуют цисты — покоящиеся стадии, устойчивые к изменению температуры, влажности и др. При окраске по Романовскому— Гимзеядро простейших окрашивается в красный, а цитоплазма—в голубой цвет. По типу питания они могут быть гетеротрофами или ауто-трофами. Многие простейшие (дизентерийная амеба, лямблии, трихомонады, лейшмании, балантидии) могут расти на питательных средах, содержащих нативные белки и аминокислоты. Для их культивирования используются также культуры клеток, куриные эмбрионы и лабораторные животные. |
|
|