Файл: Исследование действия антиоксидантов полифенольной природы на морфологию эритроцитов.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 09.11.2023
Просмотров: 66
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Изофлавоноиды обладают фенильной группой в третьем положении. Вещества содержатся в корнях Ononis arvensis (стальник полевой).
Антоцианы (керацианин, рутинозил-3-цианид в костянках Cerasusвишни; цианидин, пеларгонидин) появляются в листьях при снижении концентрации хлорофилла, тем самым обуславливают специфическую окраску плодов и листьев в растениях.
Фитоалексины на примере ресвератрола позволяют подавлять рост микрофлоры на поверхности растений. С физиологической точки зрения данные соединения оказывают возбуждающее действие на сердечную активность.
1.3 Действие полифенолов на структуру эритроцитов
Механизм воздействия соединений, включающих несколько мономерных остатков фенола, в основном выражается в качестве положительных изменений в функциональных элементах, составляющих сердечно-сосудистую систему.
Одним из эффектов, реализуемых за счёт включения полифенолов в реакции метаболизма, является запуск программы апоптоза при изменении активности белков семейства Bax/Bcl2. В тоже время происходит активизация сигнального пути фосфоинозитид-3-киназы (PI3K) – киназы AKT – мишени рапамицина млекопитающих (mTOR), характерный для коммуникации внутриклеточных структур [1].
Среди других механизмов регуляции процессов на организменном уровне, существует ряд иных клеток-мишеней, на которые воздействуют упомянутые вещества растительного происхождения. Они влияют на синтез белка VEGF при условии недостатка кислорода в среде, окружающую клетку. Схожим действием обладает фитоалексин ресвератрол, который понижает уровень синтеза данного белка. Это способствует синтезу транскрипционного фактора HIF, который отвечает за процесс эритропоэза в костном мозге [1, 16, 20].
При развитии условий для формирования окислительного стресса, вызванного сильным окислителем органической природы, негативные последствия способно нивелировать взаимодействие полифенольных соединений с эритроцитами. На примере окислителя трет-бутилгидропероксида (ТБГП) можно наблюдать увеличение концентрации перекисей и гидроперекисей липидов с 2,2 нмоль/мл до 21,7 нмоль/мл. Добавление полифенолов с разведением 10 мкМ позволило уменьшить число продуктов перекисного окисления липидов (ПОЛ) в 2 раза [2].
Присутствие большого количества гидроксильных групп в составе антиоксидантов кверцитин и кахетин позволяет реализовать защитное действие по отношению к липидам мембраны эритроцитов. Кахетин обладает свойствами, которые позволяют защитить мембрану эритроцитов не только от воздействия продуктов перекисного окисления липидов, но и препятствуют развитию перманганат-индуцированного гемолиза. Содержание кахетина и других полифенолов можно определить согласно методике Фолина-Чокальтеу [15, 21].
Меньший антиокислительный эффект показал нарингенин, обладающий свойством поляризации гидроксильных (OH-) группировок, что позволяет соединению активно участвовать в реакциях окисления и восстановления [2].
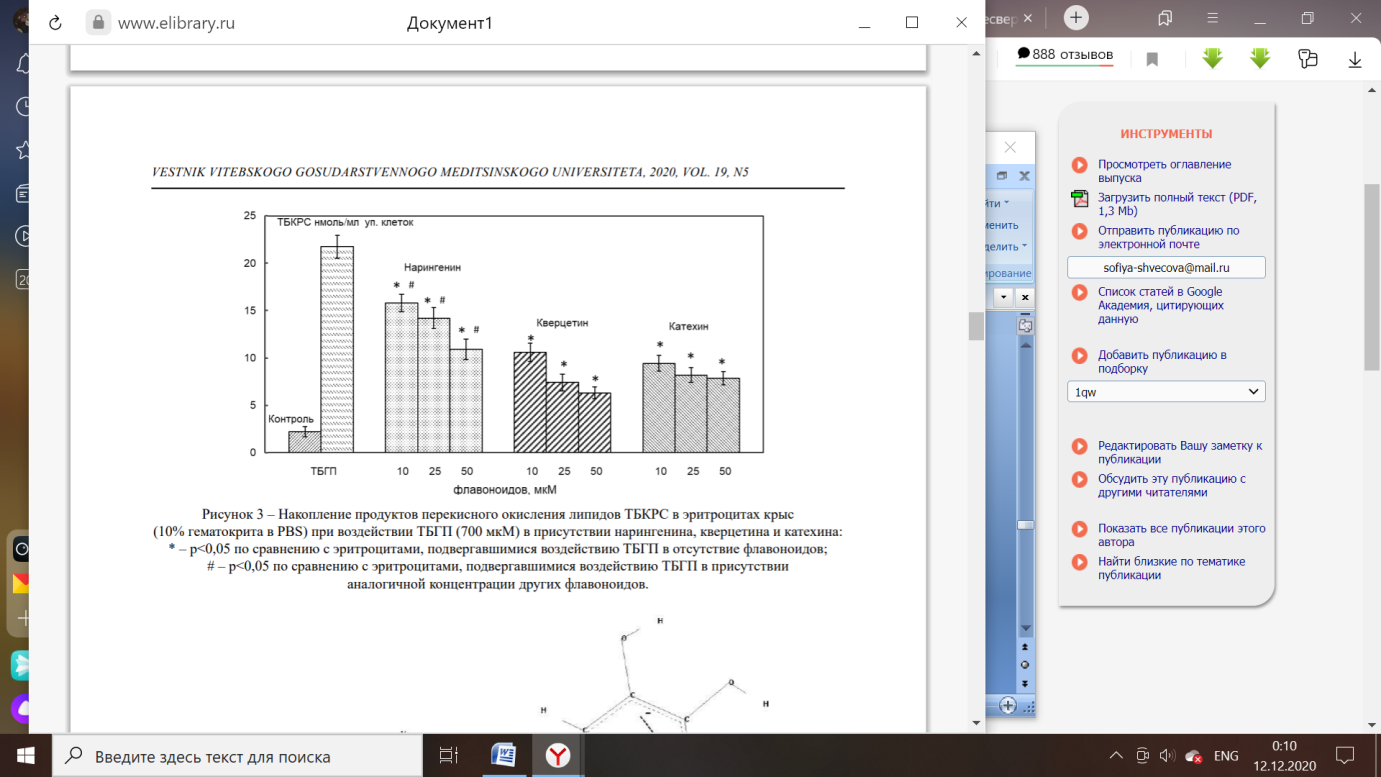
Рисунок 3 – Накопление продуктов перекисного окисления липидов в эритроцитах крыс при воздействии ТБГП (700 мкМ) в присутствии нарингенина, кверцетина и катехина [2].
При анализе распределения антиоксидантов наблюдается увеличение концентрации катехина в цитоплазме эритроцита, поскольку соединение хорошо растворимо в водных растворах. Одновременно фиксируют присутствие кверцетина в билипидном слое за счёт гидрофобных и неполярных свойств полифенола [2].
С другой стороны, внесение трет-бутилгидропероксида снижает молярную концентрацию глутатиона (GSH) в эритроците с 2,03 мкМ до 0,24 мкМ. Наличие антиоксидантов позволяет обеспечить сохранение концентрации глутатионана определённом уровне, используя субстраты окисления в составе цитоплазмы эритроцита. Эффект протекторной реакции был менее выражен чем в случае с мембранными липидами [2].
Явление асимметрии липидной структуры мембраны может развиться в эритроцитах на фоне развития сахарного диабета и повышения концентрации глюкозы в плазме крови. Антиоксидантные полифенолы растений могут взаимодействовать с гидрофильными и гидрофобными молекулами организма, обеспечивая амфифильность действия флавоноидов [4, 16].
Подобным эффектом обладает антиоксидант флоретин, основным источником которого являются листья зелёного чая и яблони. Он способен на угнетение процесса транспорта глюкозы в клетки эпителия кишечника и почек, а также ингибирование перемещения мочевины в эритроцитах и гепатоцитах. Подобным действием по отношению к глюкозе обладают полифенолы эвгенол, а также изоэугенол в составе растения семейства – герания [4, 13].
Антиоксидант флоретин способен инициировать выход флуоресцентного маркера из фосфохолиновых липосом, вызывает снижение температуры плавления дипальмитоил фосфохолина (ДПФХ). При последовательном изменении стеаринового и сфингомиелинового состава липосом молекулы полифенола распределяются между липидами разной степени упорядоченности. Это обеспечивает гетерогенность каналов, составленных липопептидом сирингомицином Е в билипидных мембранах клеток [4].
Флоретин характеризуются влияниям на термотропное поведение ДПФХ. Агент приводит к исчезновению пика на момент предперехода фосфохолинов из гель-состояния в риппл-фазу, независимо от толщины модельных мембран [4].
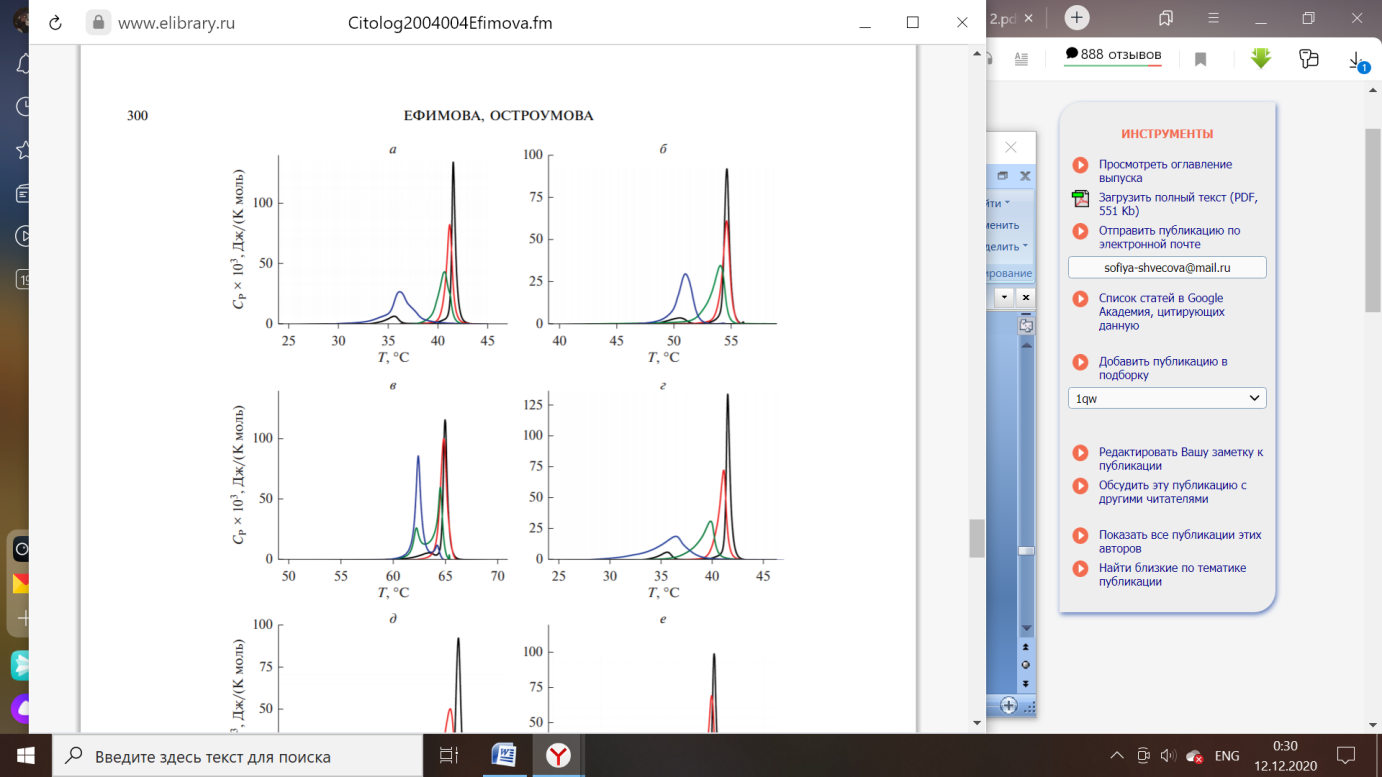
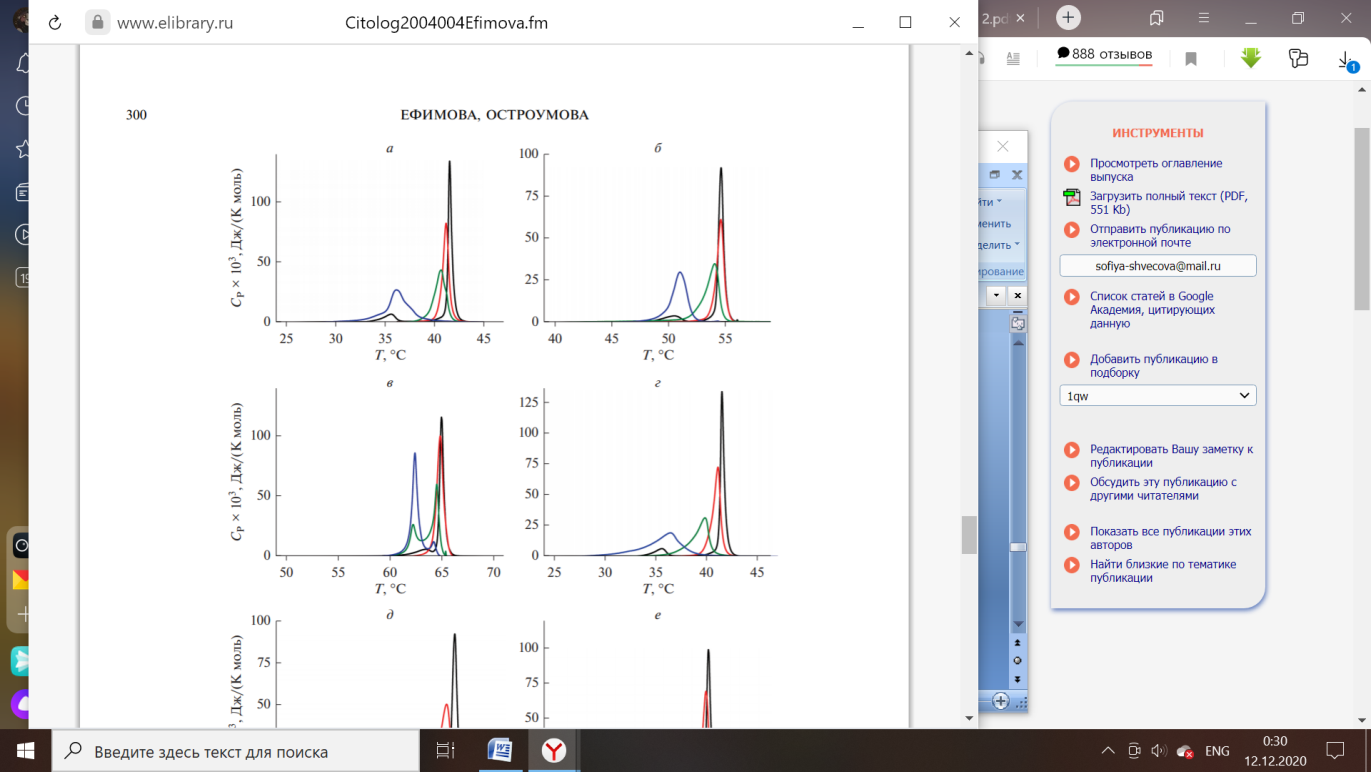
Рисунок 4 – Термограммы плавления ДПФХ (а), ДСФХ (б) и ДАФХ (в) в отсутствие агентов (чёрный) и в присутствии флоретина при соотношении липида и агента 100:1 (красный), 50:1 (зелёный) и 10:1 (синий) [4].
В ходе встраивания в структуру мембраны, растительный полифенол увеличивает площадь поверхности молекулы липида и подвижность углеводородных хвостов при падении температуры основного фазового перехода. Затем флоретин располагается в области амфифильных молекул билипидной мембраны, происходит уменьшение нарушения порядка структурных компонентов с увеличением молекулярной массы жирных кислот в составе триацилглицеридов (ТАГ), окружающих антиоксидант [4].
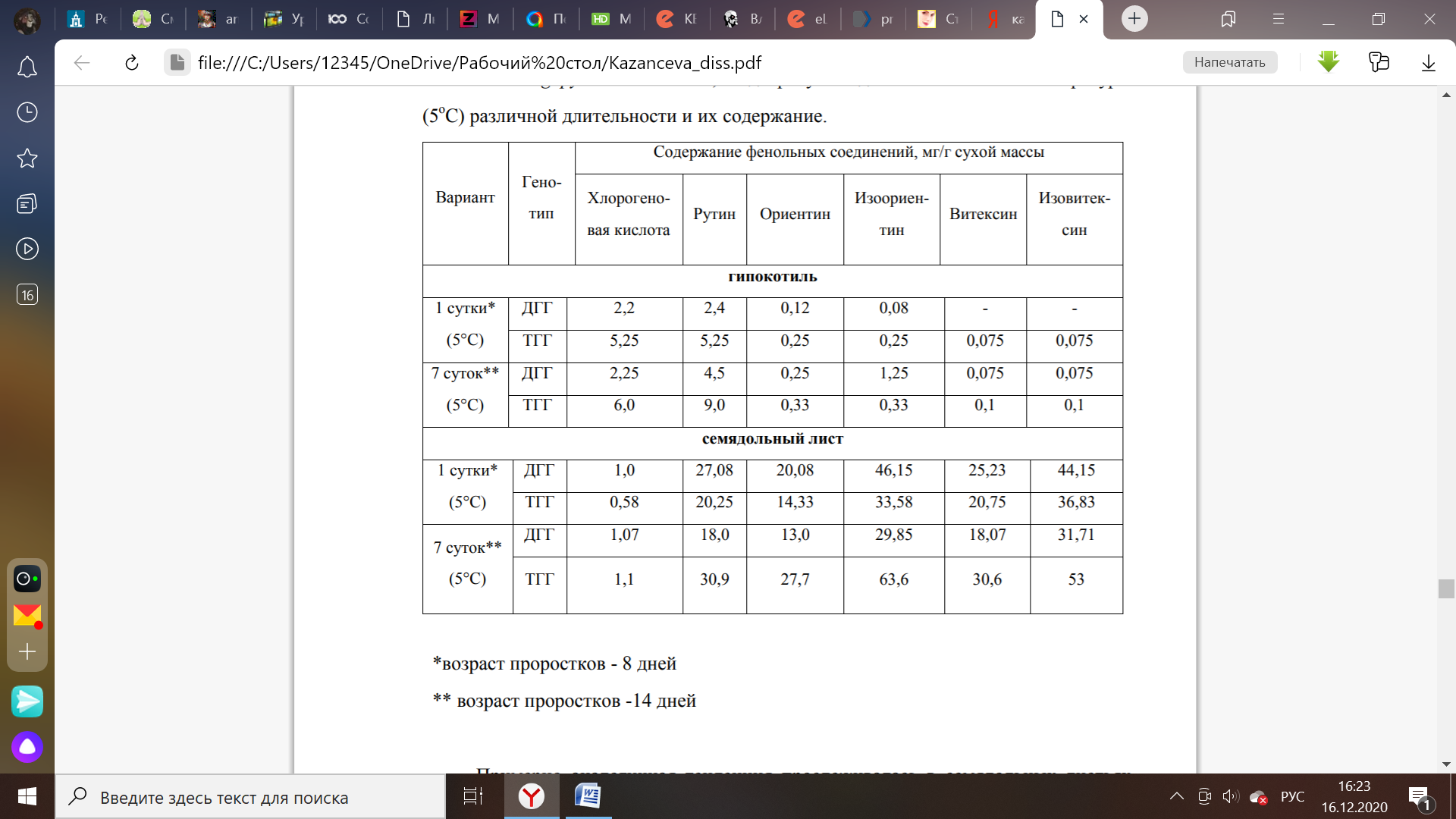
Температурный режим, в котором происходит рост и развитие растений, также оказывает влияние на концентрацию полифенольных соединений, которые накапливаются в различных компартментах организма. С уменьшением температуры проращивания отростков гречихи наблюдается увеличение содержания изоориентина на 80% по сравнению со значениями в контрольной группе растений. Подобная тенденция характерна для витексина (на 33%) при создании условий гипотермии в течение 7 суток [5].
Параллельно данный эксперимент проводили на отростках гипокоптилей гречихи с тетраплоидным набором генетического материала. При выращивании растений в условиях низкой температуры в течение 1 суток и 7 дней содержание флавоноидов в семядольных листьях было значительно выше, чем в гипокотилях: на 99% для ориентина, изоориентина, витексина, изовитексина и на 74% для рутина [5].
Кратковременное влияние гипотермии приводит к уменьшению содержания фенольных соединений в составе диплоидных образцов гречихи (Fagopyrum esculentum). У тетраплоидных отростков (ТГГ) гипокоптилей данного растения наблюдается увеличение или сохранение концентрации полифенолов на уровне, соответствующего контрольным образцам гречихи. В семядольных листьях тенденция отмечена прямо противоположная закономерность в отношении ди- (ДГГ) и тетраплоидных групп растений.
Более длительнее действие низкой температуры способствовало накоплению фенольных соединений в гипокотилях проростков ДГГ гречихи по сравнению с контролем. Подобная тенденция зафиксирована в семядольных листьях ТГГ [5].
Закономерность изменения концентрации полифенолов при кратковременном и длительном воздействии пониженной температуры может быть использовано для выделения достаточного количества биологически активных соединений для использования в условиях лабораторных экспериментов.
Таблица 2 – Основные соединения фенольного комплекса гипокотилей и семядольных листьев проростков гречихи (Fagopyrum esculentum), подвергнутых действию низкой температуры (5˚C) [5].
В общем случае действие полифенольных соединений и флавоноидов как частных представителей данной группы биологически активных веществ проявляется за счёт изменения биофизических показателей, используемые для описания состояния мембранных компонентов молекул. При добавлении полифенолов наблюдается снижение микровязкости эритроцитарной мембраны, способствующее улучшению кровотока по капиллярам сердечно-сосудистой системы исследуемого организма [22].
Добавление растительного антиоксиданта (например, кверцитина) к среде, содержащей эритроциты, вызывает потерю двояковыгнутой формы последних, наблюдается увеличение среднего диаметра на 12,05% и уменьшение толщины форменных элементов на 17%. Присутствие большого количества гидроксильных группировок в составе данного полифенола и существование π-«облака», объединяющего электроны в системе сопряжённых орбиталей, позволяет кверцитину быть донором водорода или электронов. Происходит нейтрализация свободных радикалов, гидроперекисей и перекиси водорода. Уменьшение содержания продуктов окислительных реакций позволяет снизить вероятность повреждения мембраны [22].
Также кверцетин можно идентифицировать в экстракте мякоти банана (Musa balbisiana), где в группе с другими полифенолами он обеспечивает защиту от активных форм кислорода и регуляцию сигнальных путей, запускаемых при развитии стрессовых реакций в организме [19].
Обработка эритроцитов с использованием другого флавоноида (ресвератрол) позволяет сохранить морфологические свойства на уровне, характерном для этого форменного элемента при нормальных условиях. Дискоциты не изменили среднее значение диаметра (8,15 мкм), зафиксированное до добавления ресвератрола к суспензии, в то время как толщина эритроцитов сократилась на 13%. По сравнению с кверцетином данный антиоксидант проявляет меньшую степень изменения толщины исследуемых структур при помощи атомно-силовой микроскопии. Это может служить признаком сравнения их активности как антиоксидантных соединений [22].

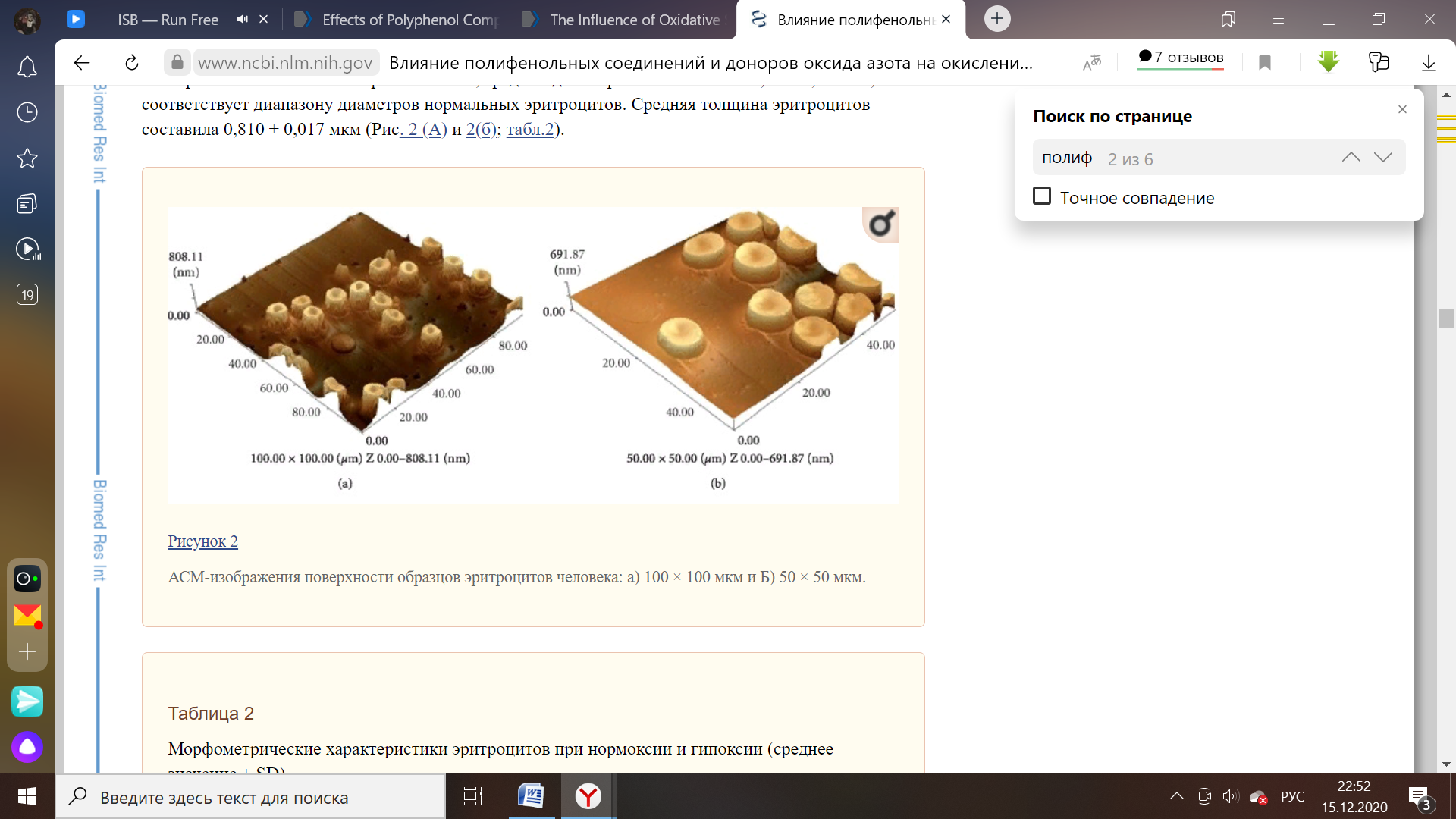
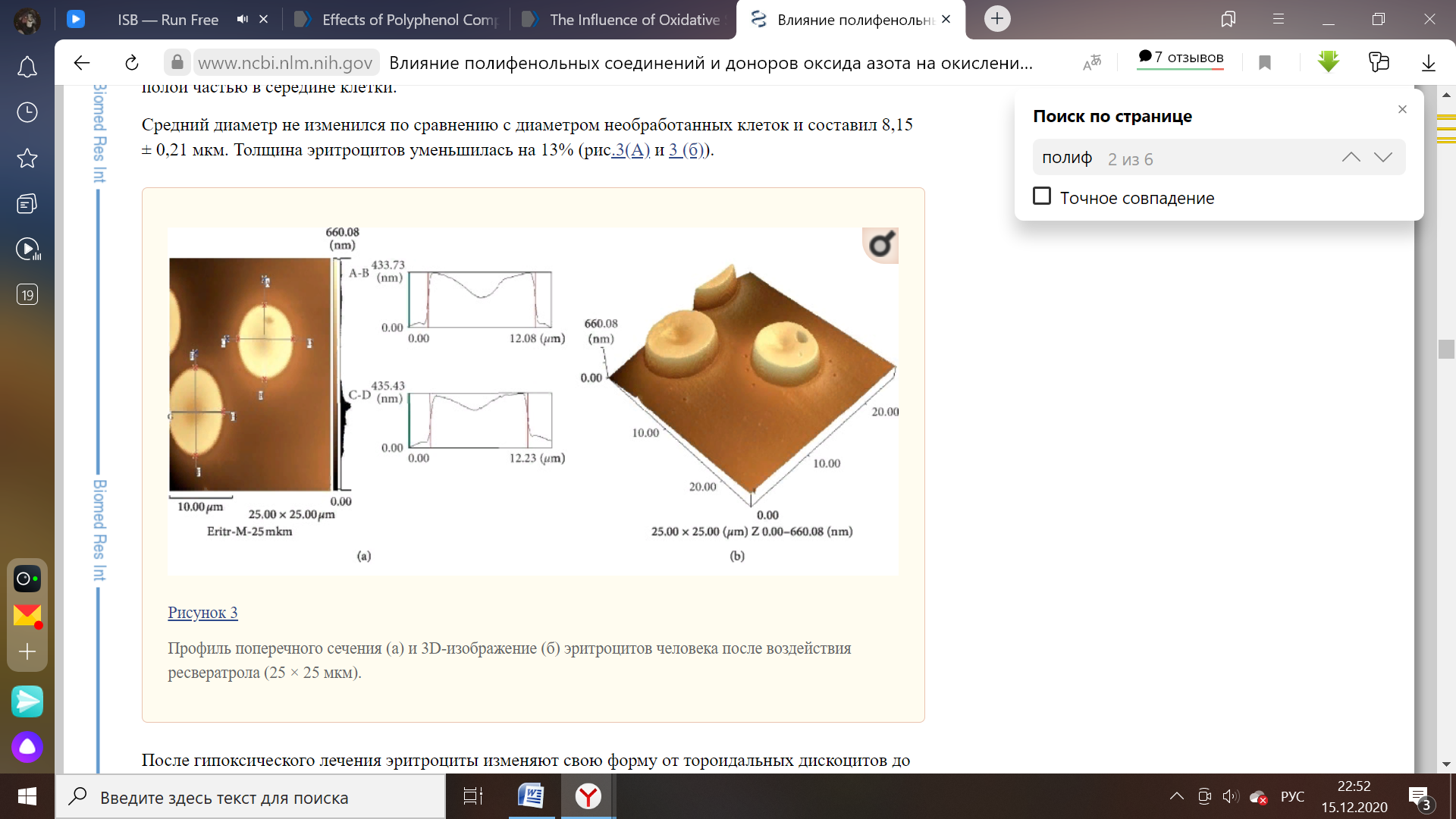
Рисунок 5 – Поверхность эритроцитов до опыта (масштаб изображения –50*50 мкм) и после добавления ресвератрола (25*25 мкм) [22].
Помимо описанных свойств полифенол способен реагировать с активными формами кислорода. При сравнении степени проявления данного свойства у других флавоноидов наиболее интенсивным эффектом обладает кофейная кислота в составе листьев картофеля [19, 26].
Гликозидная форма ресвератрола может образовывать конъюгированные соединения, обладающие хорошей растворимостью в водных растворах [23].
Особое значение приобретает возможность данного антиоксиданта оказывать влияние на конформационное состояние молекулы гемипорфирина – основного пигмента в составе эритроцитов. Изменение структуры металлопротеина коррелирует со способностью гемоглобина связывать молекулы кислорода [23].
Реакции окисления липидов и белковых молекул в составе эритроцитов имеют регуляцию за счёт ферментативных систем клеток и при добавлении антиоксидантных соединений, в том числе растительного происхождения.
Флавоноид ресвератрол образует пероксидные радикалы, обладающие свойствами стабильности в условиях резонанса частот в ходе анализа рамановских спектров. Данные радикалы образуются по принципу углеродной одинарной связи. Одновременно наблюдается снижение уровня перекиси водорода при добавлении ресвератрола, поскольку соединение принимает электроны от радикалов, обладающих реактогенными неспаренными электронами [23].
Использование лекарственных препаратов может способствовать увеличению устойчивости мембраны эритроцитов, а также обуславливать её чувствительность к разрушению целостности мембранных липидов и белков при введении антагониста данного препарата [27].

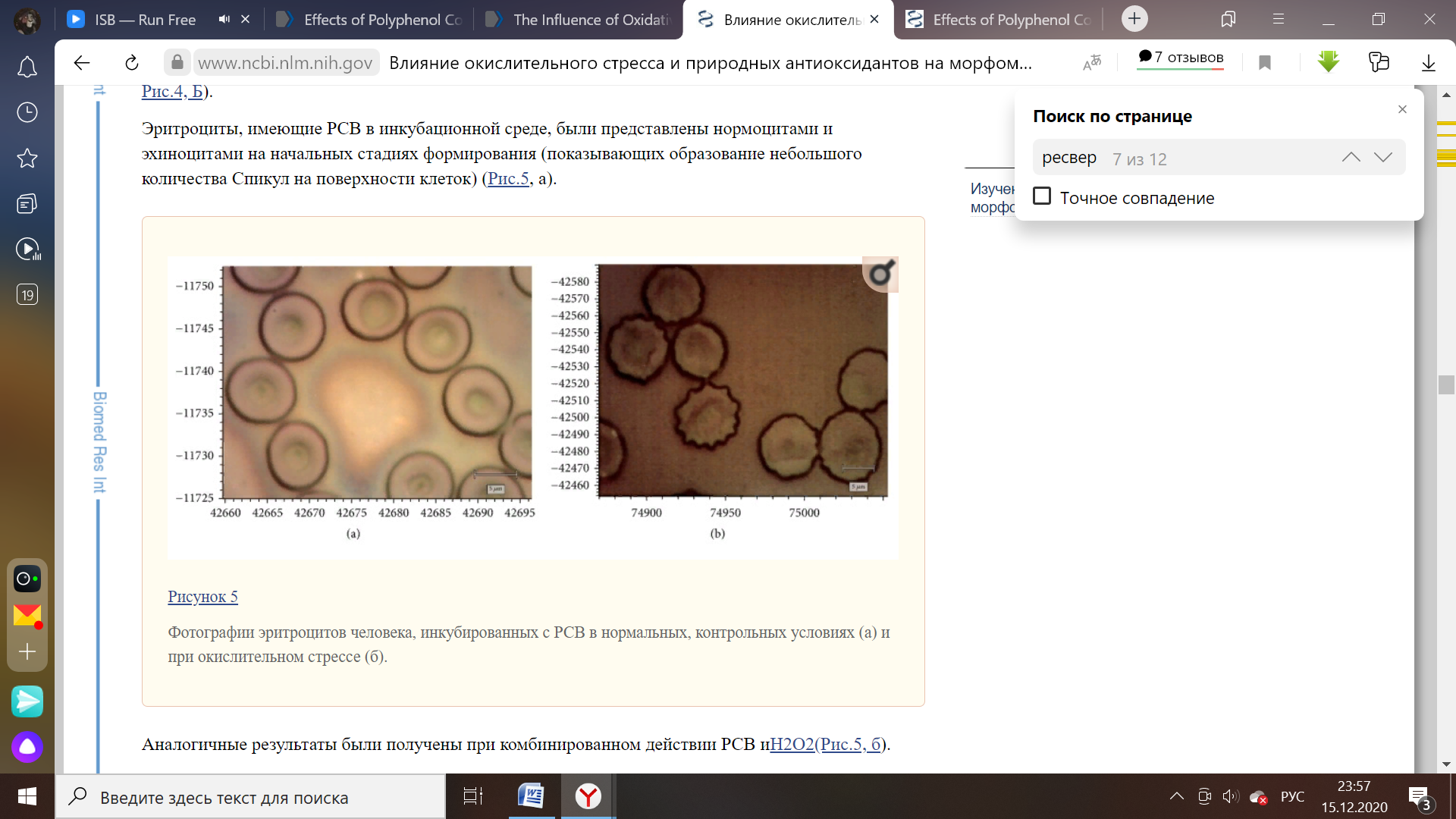
Рисунок 6 – Эритроциты человека в нормальных условиях и при формировании окислительного стресса [23].
Некоторые условия окружающей среды (свет, температура или наличие кислорода в среде) могут влиять на физико-химическую стабильность полифенолов. Для преодоления этих ограничений используют различные модели загрузки полифенолов в наночастицы альбумина, в том числе адресная доставка действующего вещества характерна для полифенола куркумин [27, 29, 30].