ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.12.2021
Просмотров: 269
Скачиваний: 4
ТЕМА 9. МЕХАНІКА КЛІТИННОЇ МЕМБРАНИ
9.1. Молекулярна структура клітинної мембрани
Біологічна мембрана - одна з найдавніших компонент живих клітин, структура якої однакова в основних рисах, незалежно від того чи це мембрана ядра, ендоплазматичного ретикулуму, чи зовнішня оболонка клітини. При цьому мембрана виконує складні функції: селективного бар'єра для різних речовин, діелектричного прошарку, поверхні розділу, матриксу, а також структури, що визначає форму клітини.
Щоб зрозуміти механічні властивості клітинної мембрани, необхідно розглянути її молекулярний склад. У 1972 році Зінгер і Ніколсон сформулювали рідкомозаїчну модель клітинної мембрани.
Приблизно на 40 % клітинна мембрана складається з ліпідів L, які утворюють бімолекулярний шар (рис.9.1).
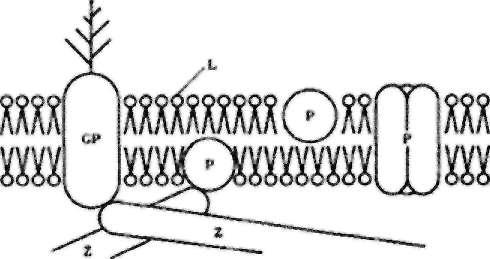
Рис.9.1. Структура клітинної мембрани.
Ліпідний склад мембрани (фосфоліпіди, холестерин, жирні кислоти, тригліцериди та ін.) змінюється залежно від типу і функцій клітини, а також фаз її життєвого циклу. В результаті так званого "фліп-флопу", можливий обмін шарами мембрани. In vivo можливий обмін ліпідами із зовнішнім середовищем, наприклад, плазмою крові.
В ліпідний матрикс вкраплені білки Р і функціональні білкові комплекси. Частину з цих включень становлять глікопротещи GP, які виступають в замембранний простір і утворюють "трухляву" поверхню клітини. Ці виступи є носіями електростатичного заряду і відіграють одночасно важливу роль в імунній системі клітини. Білки є основним компонентом, який відповідає за вибіркову проникливість клітинних мембран. Вони утворюють систему селективних "nop" або функціонують як іонні помпи і регулюють, наприклад, електрохімічну систему збудження нервової клітини.
Біля зовнішньої поверхні знаходяться адсорбовані молекули і молекули, зв'язані з мембраною в результаті імунних реакцій. З внутрішньої сторони мембрани знаходяться білки, які утворюють цитоскелет. Зовнішні і внутрішні позамембранні Z компоненти обмежують вільне переміщення білків у мембрані.
На основі розглянутої молекулярної структури клітинної мембрани можна зробити ряд якісних передбачень:
1. Товщина мембрани (6... 10 нм) практично не залежить від механічних навантажень, а визначається лише молекулярним складом.
-
Віддаль між молекулами мембрани, а також площі, які займають
молекули, визначаються електростатичними і вандерваальсовими силами. -
Різні молекули мембрани можна розглядати як циліндри або конуси.
Мембрана може змінювати свою кривизну при зміні площі одного з моношарів
(за аналогією з біметалева пластиною).
9.2. В'язкопружні властивості клітинної мембрани
Вивчення механічних властивостей мембран клітин і особливо кров'яних клітин є актуальним завданням сучасної біомеханіки. Кров людини містить кров'яні клітини декількох типів, але на її властивості найбільший вплив мають еритроцити. Це зумовлено їх високим об'ємним вмістом (близько 45 %) і концентрацією (близько 5' 10 мм'3). Тому вивчення механічних властивостей еритроцитів має важливе значення для пояснення руху крові в судинах (особливо малого діаметра), швидкості осідання еритроцитів, процесів руйнування еритроцитів тощо.
У недеформованому стані еритроцити мають форму дуже гнучких двояковигнутих дисків рис.9.2.
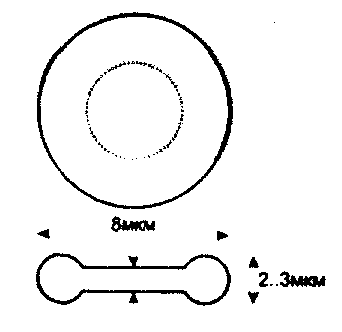
Іикм
Рис.9,2. Схематичне зображення недеформованого еритроцита.
Форма еритроцитів має важливе значення для забезпечений їх основної функції - транспортування кисню та Інших речовин. При поперечному розрізі еритроцит нагадує форму гантелі. Діаметр еритроцита дорівнює близько 8,5 мкм, товщина - 2,2 мкм, а об'єм - близько 90 мкм . Загальна поверхня еритроцитів досягає 3000 м2. Якщо б еритроцити мали форму кулі, то загальна площа їхньої поверхні була б на 20% меншою і значно б сповільнилася швидкість газообмінних процесів.
Еритроцити складаються з тонкої мембрани і рідкого наповнення, яким є майже насичений розчин гемоглобіну (густина цільного еритроцита дорівнює 1,08103кг/м3).
Близько 90% сухої речовини еритроцитів становить гемоглобін. Він знаходиться всередині еритроцита, це забезпечує зменшення в'язкості крові. Одночасний гемоліз (руйнування мембрани еритроцитів з виходом гемоглобіну в плазму) всіх еритроцитів призвів би до збільшення в'язкості крові в декілька разів і це різко ускладнило би роботу серцево-судинної системи.
Товщина мембрани еритроцита становить 7,5 нм і її маса не перевищує З % від маси клітини. Оскільки товщина мембрани є значно меншою від лінійних розмірів еритроцита і радіусів кривизни (мінімальний радіус кривизни -близько 1 мкм), для аналізу його властивостей можна використати теорію тонких оболонок. Згідно з цією теорією, опір, який чинить мембрана навантаженням розтягу, значно перевищує опір мембрани до навантажень згинання. Висока гнучкість мембрани еритроцитів і їх рідке наповнення, яке
-58-
чинить при зміні форми лише в'язкий, а не пружний опір, забезпечують еритроцитам добрі деформаційні властивості. Важливу роль у цьому відіграє також форма еритроцитів.
Подібно до інших несферичних тіл еритроцити можуть набувати найрізноманітнішої форми без зміни об'єму і площі поверхні, тобто без розтягання мембрани. Це пояснюється високою жорсткістю мембрани при розтяганні (модуль Юнга має порядок 106Н/м2 і практично не змінюється при деформації) та низькою межею міцності при розтяганні. Відомо, що сферичне тіло займає максимальний об'єм при даній площі поверхні. Тому при деформуванні такого тіла площа його поверхні повинна збільшуватись. У випадку еритроцитів це неможливо, тому при розбуханні еритроцита (наприклад, при зменшенні осмотичного тиску плазми крові) він спочатку набуває форми сфери, а потім - руйнується мембрана.
Еритроцити змінюють свою форму в потоці рідини з градієнтом швидкості. При напруженнях зсуву до 10 Н/м деформація еритроцитів є незначною. Збільшення тангенціальних напружень до 200 Н/м2 призводить до руйнування клітин. Середня тривалість життя еритроцита становить 120 днів. Із старінням мембрана еритроцитів стає крихкою і при деформуванні руйнується. Найчастіше це відбувається у дрібних судинах (там найбільші зсувні напруження) і турбулентних потоках крові.
Для вимірювання механічних властивостей клітинної мембрани використовують різні підходи - від молекулярних методів до феноменологічних досліджень деформованості клітини під дією заданого навантаження.
Феноменологічні методи вимірювання в'язкопружних властивостей клітинної мембрани зводяться до певної зміни форми клітини зовнішніми силами. На рис.9.3 показані деякі розроблені за останній час методи.
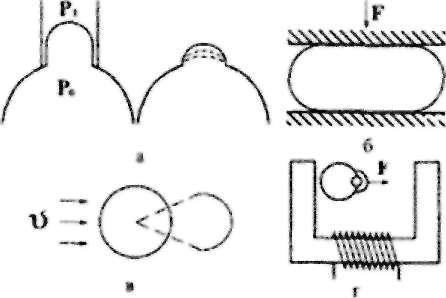
Рис.9.3. Методи вимірювання в'язкопружних арактеристик клітинної мембрани.
Якщо засмоктати еритроцит у капіляр (піпетку) певного діаметра, то за прикладеною різницею тисків (ро - рі) і радіусами кривизни частин клітини можна визначити механічні параметри еритроцита. Для зменшення впливу контакту капіляра на мембрану оцінюють швидкість релаксації клітини після звільнення ЇЇ з капіляра (рис.9.3,а).
Інші методи полягають у деформуванні еритроцитів у потоці рідини (рис.9.3,б), між двома плоскопаралельними пластинками (рис.9.3,в) або в
-59-
результаті руху введених у клітину феромагнітних частинок під дією зовнішнього магнітного поля (рис.9.3,г).
Результати подібних досліджень показують, що на відміну від гумової мембрани, біологічна мембрана практично не розтягується. Середні поверхневі деформації клітини становлять лише близько 0,01 %. Межа міцності на розтягання мембрани дуже низька. Тому осмотичне набухання клітини можливе лише до того моменту, поки вона не досягне ідеальної кулеподібної форми. Розрив мембрани настає вже при тиску в клітині менше ніж 100 Па.
Клітинна мембрана легко піддається деформації паралельного зсуву. В потоці з градієнтом швидкості за видимими мітками на мембрані можна спостерігати обертання мембрани навколо свого наповнення. Це явище називається феномен гусениці танка.
Механічні властивості клітинної мембрани еритроцитів значною мірою визначають форму клітини і динаміку клітинного руху в кровоносному руслі.