Файл: Курсовая работа научный регистрации по журналу учета курсовых работ.docx
Добавлен: 11.01.2024
Просмотров: 60
Скачиваний: 2
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Рис. 2. Кривая цветения травянистых видов под пологом дубравы (липо–дубняк. лесостепь) (по: Горышина, 1969): А, 1 – число цветущих видов; Б, 2–относительная освещенность (%) на уровне травяного покрова.
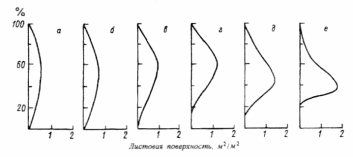
Рис. 3. Распределение листовой поверхности по высоте у лабазниковых (Filipendula camtschatica) травостоев в расчете на участок площадью 1 м2, Камчатка (по: Степанова и др., 1981): а–15 июня, 6–23 июля, в –6 июля, г – 15 июля, д–28 июля, е –9 августа.
Сезонная изменчивость сообществ неодинакова. Она определяется со– ставом сообществ и особенностями изменения экологических факторов в разных климатических зонах и экотопах. В упомянутом лабазниковом со– обществе крупные травы образуют при полном своем развитии сплошной листовой полог. Под ним возникает крайний дефицит света, препятствующий произрастанию низкорослых растений. Листовая поверхность в течение сезона «поднимается» вверх. В высокотравном луговом сообществе в степной зоне с увеличением размеров растений также наблюдается такой подъем. Однако в целом листовая поверхность оказывается более или менее равномерно распределенной по высоте, поскольку здесь не образуется плотного полога, перехватывающего свет (рис. 4).

Рис. 4. Изменение вертикального распределения относительной листовой поверхности (ОЛП) в высокотравном луговом сообществе, степная зона, дно балки (по: Утехин, 1977).
1 – начало июня, 2– начало августа.
В широколиственных листопадных лесах весной при обилии света под необлиственным еще древостоем развиваются синузии эфемероидов, часто образующие сплошной ковер, но к середине лета в надземной своей части полностью исчезающие. В сомкнутых вечнозеленых еловых лесах таежной зоны из–за ограничения светопого довольствия такие возможности для эфемероидов отсутствуют. Сезонная смена синузий здесь не выражена. Под пологом березовых же лесов в тайге синузия эфемероидов успевает сформироваться до полного развития листвы березы, правда видовой состав синузии беден и представлен в основном ветреницей (Апетопа nemorosa). Впрочем, в вегетативном состоянии многие особи ветреницы сохраняются в течение всего лета (Ипатов, 1997).
В степи после образования основной массы в весенний и раннелетний период ее уровень мало меняется; Ограничивают образование фитомассы высокие температуры, а главное, недостаточное снабжение растений водой. На субальпийских лугах, в условиях обеспеченности влагой динамика фитомассы следует за изменением температурного режима (рис.5).

Рис. 5. Сезонное изменение зеленой массы в разных сообществах (Гордеева, 1965; Маилов, 1971; Коваленко и др., 1973): 1 – тонковатополынно–типчаково–ковылковое, 2–тонковатополыннотырсиковое (степные сообщества, Центр. Казахстан); 3–злаково–бобово–разнотравный послелесной субальпийский луг, заповедный; 4 – красно–овсяницевый луг, 5–белоусовый луг – субальпийские луга в Карпатах.
Своеобразный ритм развития имеют пустынные сообщества. Во многих пустынных сообществах однолетние растения создают основные аспекты (внешний вид) и производят большую часть фитомассы. В сезонном ритме сообществ выделяются три группы видов: ранневесенние эфемеры, поздневесенние эфемеры и летние однолетники (Нечаева. Василевская, 1945). Они различаются строением, позволяющим растениям адаптироваться к режиму влажности, тепловому режиму в разное время вегетационного периода. Ранневесенние эфемеры (Ceratocephalus falcatns. Meniocus linifolius, Streptoloma desciforum) прорастают в ноябре, цветут в начале марта и засыхают в середине апреля. Они развиваются в период с достаточным количеством влаги и сравнительно невысокими для пустыни температурами и имеют мезоморфное строение. Поздневесенние эфемеры (Lappula seniglabra, Нурссоит pendulum, Malcolmia grandiflora. Isatis emarginaia, Senecio subdeniatus, Centaurea gyzensis, Delphinium camptocarpum) прорастают в декабре, цветут в апреле, засыхают к середине мая. Ксероморфные черты (утолщенная пластинка листа, столбчатая ткань с верхней и с нижней стороны) позволяют им развиваться в условиях большего дефицита влаги и более высоких температур. Продолжительность вегетации летних однолетников больше, чем у эфемеров, и проходит в другие сроки: прорастают они в марте–апреле и завершают развитие осенью (в конце сентября – начале октября). Эти виды – представители семейств Chenopodiaceae и Euphorbiaceae (Chrozophora sabulosa, Euphorbia cheirokpis, Salsola pellucida, Horamnovia uhcina. Agnophyllum latifohum, A.minus, Londesia enantha, Kochia schrenkiana. Airiplex dimorphostegia), развиваются в жестких условиях ограниченного водного довольствия и высоких температур. Устойчивость этих растений к засухе обеспечивается тем, что в их стеблях и листьях развита крупноклеточная водозапасающая ткань, в тканях накапливаются хлориды и сульфаты, благодаря этому клеточный сок имеет повышенное осмотическое давление. У растений семейства Euphorbtaceae имеются млечники, заполненные слизистыми веществами, смолами, маслами. Разумеется, в сообществах участвуют и другие виды, многолетники, как монокарпики, так и поликарпики. В пустыне сезонная динамика отличается глубокими и резкими изменениями. В описанных пустынных сообществах наблюдаются два пика вегетации: летний и осенне–весенний. В некоторых экотопах сезонный ритм может в основном зависеть от биологических особенностей самих растений. В тугайных травяных сообществах, монодоминантных, прирост фитомассы определяется ритмом развития доминирующих видов. Прирост фитомассы прекращается по достижении доминантами предельных размеров. Период же роста у разных видов неодинаков. Например, у Typha minima он заканчивается в конце июня, а у Phragmites australis продолжается непрерывно в течение вегетационного периода (
Ипатов, 1997).
Последовательность сезонных изменений у одного и того же сообще– ства из года в год повторяется, но сроки прохождения фаз и отдельные частности различаются.
2.2 Флуктуация
Если условия произрастания видов в сообществе заметно меняются от года к году, может наблюдаться разногодичная изменчивость – флуктуации. Продолжительность цикла флуктуаций не превышает 10 лет. Например, урожайность пойменного луга в долине реки Белой в разные года изменяется в 10 раз. Флуктуации фитоценозов у северной границы степей приводят к тому, что границы степи во влажный год может сместиться на юг, а в сухой – на север. В экстремальные годы граница может сместиться на 100 км.
Разногодичная изменчивость, или флуктуации, – это изменения фитоценоза от года к году. Они чаще всего связаны с тем, что условия существования растений в разные годы не одинаковы, причем иногда различаются в сильной степени (например, в некоторые годы выпадает много осадков, в другие – мало).
Разногодичная изменчивость фитоценозов хорошо выражена на пойменных лугах, заливаемых весной при разливе рек. На этих лугах в разные годы сильно разрастаются и становятся доминирующими то одни растения, то другие. Количественное соотношение разных видов растений год от года сильно меняется, хотя общий видовой состав остается неизменным. Разногодичные изменения в данном случае обусловлены тем, что отдельные годы сильно различаются между собой по продолжительности весеннего паводка. В годы с продолжительным паводком сильно разрастаются более влаголюбивые растения (например, лисохвост луговой), в годы с кратковременным паводком – менее влаголюбивые (например, клевер луговой).
Разногодичные изменения фитоценозов хорошо выражены и в степях. Здесь в некоторые года очень обильно цветут ковыли и степь летом напоминает седое море, которое колышется от ветра. В другие же годы ковыли цветут, напротив, довольно слабо и не дают сплошного фона. Отмеченное явление объясняется тем, что в разные годы выпадает то больше, то меньше осадков. В условиях степи на фоне общего дефицита влаги это очень важно.
Если от года к году условия жизни растений меняются плавно, то разногодичная изменчивость фитоценозов выражена слабо. Это имеет место, например, в таежных хвойных лесах, где растения всегда обеспечены влагой. Фитоценозы тайги от одного года к другому меняются сравнительно мало.
При разногодичных изменениях фитоценозов видовой состав растений остается прежним. Изменяются только господствующие растения, численность цветущих экземпляров и т.д.
Происходят как бы колебания около определенного «среднего» уровня. Они не приводят к перестройке фитоценоза и смене его каким–либо другим.
Возрастная изменчивость наблюдается только в лесных фитоценозах, причем таких, где деревья более или менее сходны по возрасту (древостой бывает одновозрастным обычно тогда, когда лес сформировался на сплошной вырубке, пожарище, заброшенной пашне и т.д.). На протяжении жизни одного поколения деревьев, образующих древостой, происходят существенные изменения в нижних ярусах лесного фитоценоза – подлеске, травяно–кустарничковом ярусе, мохово–лишайниковом покрове. Это хорошо заметно, например, в еловых лесах таежной полосы. В молодом, очень густом ельнике из–за сильного затенения на почве не развиваются никакие растения. В ельнике более старшего возраста, где света под пологом становится больше, появляется сначала моховой покров, а затем отдельные травы и кустарнички. В старом ельнике обычно хорошо развит как моховой, так и травяно–кустарничковый покров Возрастные изменения в нижних ярусах лесных фитоценозов обусловлены тем, что с возрастом леса увеличивается освещенность под пологом деревьев, ослабевает напряженность корневой конкуренции в почве за влагу и питательные вещества и т.д. Возрастные изменения происходят также и в древесном ярусе (Прокопьева, 2009).
По мере увеличения возраста деревьев они становятся крупнее, число их на единице площади уменьшается и т.д.
Возрастные изменения лесных фитоценозов обычно носят циклический характер. Достигнув старого возраста, древесный ярус распадается, отмирает. Образуется открытое пространство, на котором вновь начинает формироваться лесной фитоценоз. Он снова проходит в своем развитии различные возрастные стадии (Прокопьева, 2009).
Сами флуктуации изменяются во времени в том смысле, что полного возврата к прошлому состоянию не происходит. Это объясняется тем, что движущая сила разногодичных изменений – погодные условия одно– временно колеблются в разных ритмах. На краткосрочные колебания накладываются долгосрочные ритмы, охватывающие большое число лет. Экотоп, в частности климат, имеет и длительные тенденции изменения, вызывающие соответствующие сдвиги в сообществах. Изменяющаяся растительность меняет и биотоп, что также приводит к направленным изменениям в сообществах. Если флуктуации трактовать как колебания параметров сообщества около средней периодической характеристики и принять во внимание, что средняя по указанным причинам имеет тренд, то становится очевидным, что полной цикличности во флуктуационной изменчивости и не может быть. Иллюстрирует высказанные соображения схема наложения всех изменений друг на друга (рис.6).
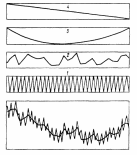
Рис.6. Схема совмещения (внизу) сезонных изменений (1) , краткосрочных флуктуации (2), длительных флуктуации (3) и направленного изменения (4) растительного сообщества.
Подобная схеме картина обнаружена в исследовании В. А. Зайковой
(1980). В течение 15 лет она наблюдала изменения луговой растительности в Карелии (рис.7). За этот период температура, осадки, баланс влаги, естественно, не оставались постоянными.
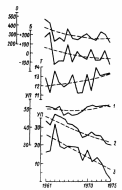
Рис. 7. Флуктуации и тенденция изменения параметров среды луговых сообществ с господством: Festuca praitnsis (1), Agrostis tennis (2), Nardus siricta (3) (Средняя тайга, Карелия) (по материалам: Зайкова, 1980): О – осадки, мм; Б – баланс влаги; X–средняя суточная температура воздуха за вегетационный период (20 мая – 30 сентября), У П – увлажнение и плодородие, оцененные по растительности, в условных единицах.
Заключение
Главнейшими особенностями фитоценоза являются взаимодействие между образующими его растениями, с одной стороны, и взаимодействие между растениями и окружающей средой – с другой. Влияние растений друг на друга имеет место только тогда, когда они более или менее сближены, соприкасаются своими надземными или подземными органами. Совокупность отдельно стоящих растений, не оказывающих влияния друг на друга, нельзя назвать фитоценозом (Прокопьева, 2009).
Формы влияния одних растений на другие разнообразны. Они были подробно рассмотрены нами ранее. Однако не все эти формы имеют одинаковое значение в жизни растительных сообществ.
Совместная жизнь растений в фитоценозе, когда они в той или иной степени влияют друг на друга, накладывает глубокий отпечаток на их внешний облик. Это особенно заметно в лесных фитоценозах. Деревья, образующие лес, сильно отличаются по внешнему виду от одиночных деревьев, которые выросли на открытом месте. В лесу деревья более или менее высокие, кроны их узкие, высоко подняты над землей. Одиночные деревья значительно ниже, кроны их широкие, низко опущенные (Василевич, 1983)
Результаты влияния растений друг на друга хорошо заметны также в травяных фитоценозах, например на лугах. Здесь растения имеют меньшие размеры, чем при одиночном произрастании, менее обильно цветут и плодоносят, а некоторые и вовсе не цветут. В фитоценозах любого типа растения взаимодействуют между собой и это сказывается на их внешнем облике и жизненном состоянии (