ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 29.10.2023
Просмотров: 698
Скачиваний: 1
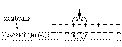 Под влиянием раздражителя (П) на мембране волокна возникает возбуждение - ПД (на рисунке снаружи "-", внутри "+" - показана фаза реверсполяризации ПД - максимум развития пика). Между возбужденными и невозбуждёнными (снаружи "+" внутри "-" - потенциал покоя) участками мембраны волокна возникает разность потенциалов, ΔU; снаружи и внутри эти участки соединены проводящей среде (цитоплазма и межклеточная жидкость) между этими участками мембраны возникают местные елекрични токи, направленные от "+" до "-", действуют на мембрану с внешней и внутренней поверхности. Эти токи являются раздражителями для мембраны. Необходимо оценить параметры этих токов как раздражителя:1. Направление - с рисунка видно, что токи имеют выходной (катодный) направление в невозбуждённых участках мембраны. Итак, здесь будет возникать деполяризация мембраны. Если она дойдет до Екр, то возникнет ПД.2. Сила - численно сила тока в данном случае равна разности потенциалов между возбужденными и невозбуждёнными участками мембраны и эта сила соответствует амплитуде ПД. Порог силы соответствует величине порогового потенциала (ΔV). Амплитуда ПД нервного волокна составляет 100-120 мВ, порог деполяризации - 15-20 мВ. Соответственно, сила раздражителя в несколько раз (6-8) больше пороговой. Отношение амплитуды ПД к порогу деполяризации называется коэффициент надежности и показывает, во сколько раз сила местного тока как раздражителя больше пороговую силу.3. Время действия раздражителя - отвечает продолжительности ПД и в несколько раз больше порогового.4. Скорость увеличения силы - соответствует скорости пика ПД, почти соответствует скорости увеличения силы при действии прямоугольных импульсов электрического тока - намного выше порога.Итак, на воспаленную участок мембраны нервного волокна действует катодный электрический ток, сила, время действия и скорость увеличения силы, которого высшие порога - этот ток вызовет деполяризацию мембраны к Екр вызовет ПД на мембране невозбужденном участка.Далее процесс повторяется ПД распространяется вдоль всей мембраны нервного волокна.М
Под влиянием раздражителя (П) на мембране волокна возникает возбуждение - ПД (на рисунке снаружи "-", внутри "+" - показана фаза реверсполяризации ПД - максимум развития пика). Между возбужденными и невозбуждёнными (снаружи "+" внутри "-" - потенциал покоя) участками мембраны волокна возникает разность потенциалов, ΔU; снаружи и внутри эти участки соединены проводящей среде (цитоплазма и межклеточная жидкость) между этими участками мембраны возникают местные елекрични токи, направленные от "+" до "-", действуют на мембрану с внешней и внутренней поверхности. Эти токи являются раздражителями для мембраны. Необходимо оценить параметры этих токов как раздражителя:1. Направление - с рисунка видно, что токи имеют выходной (катодный) направление в невозбуждённых участках мембраны. Итак, здесь будет возникать деполяризация мембраны. Если она дойдет до Екр, то возникнет ПД.2. Сила - численно сила тока в данном случае равна разности потенциалов между возбужденными и невозбуждёнными участками мембраны и эта сила соответствует амплитуде ПД. Порог силы соответствует величине порогового потенциала (ΔV). Амплитуда ПД нервного волокна составляет 100-120 мВ, порог деполяризации - 15-20 мВ. Соответственно, сила раздражителя в несколько раз (6-8) больше пороговой. Отношение амплитуды ПД к порогу деполяризации называется коэффициент надежности и показывает, во сколько раз сила местного тока как раздражителя больше пороговую силу.3. Время действия раздражителя - отвечает продолжительности ПД и в несколько раз больше порогового.4. Скорость увеличения силы - соответствует скорости пика ПД, почти соответствует скорости увеличения силы при действии прямоугольных импульсов электрического тока - намного выше порога.Итак, на воспаленную участок мембраны нервного волокна действует катодный электрический ток, сила, время действия и скорость увеличения силы, которого высшие порога - этот ток вызовет деполяризацию мембраны к Екр вызовет ПД на мембране невозбужденном участка.Далее процесс повторяется ПД распространяется вдоль всей мембраны нервного волокна.М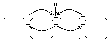 еханизм проведения возбуждения по миелиновой нервным волокнам принципиально не отличается от механизма проведения по безмиелиновых волокнах: под воздействием раздражителя (П) в одном из перехватов Ранвье возникает ПД - на мембране перезарядка между этим (возбужденным) и соседними (невозбуждёнными) перехватами Ранвье ' есть возникает разность потенциалов ΔU; они соединены проводящей средой возникают местные токи (от "+" до "-"). Эти токи в области невозбуждённых перехватов имеют исходное направление; их сила (амплитуда ПД), длительность (продолжительность ПД), скорость увеличения силы (скорость увеличения пике ПД) сверхпороговые на мембране невозбуждённого перехвата Ранвье возникает деполяризация, которая достигает Екр возникает ПД.
еханизм проведения возбуждения по миелиновой нервным волокнам принципиально не отличается от механизма проведения по безмиелиновых волокнах: под воздействием раздражителя (П) в одном из перехватов Ранвье возникает ПД - на мембране перезарядка между этим (возбужденным) и соседними (невозбуждёнными) перехватами Ранвье ' есть возникает разность потенциалов ΔU; они соединены проводящей средой возникают местные токи (от "+" до "-"). Эти токи в области невозбуждённых перехватов имеют исходное направление; их сила (амплитуда ПД), длительность (продолжительность ПД), скорость увеличения силы (скорость увеличения пике ПД) сверхпороговые на мембране невозбуждённого перехвата Ранвье возникает деполяризация, которая достигает Екр возникает ПД.
Существенное отличие распространение ПД по миелиновой волокнам заключается в том, что в них местные токи возникают не между соседними участками мембраны, а между соседними перехватами Ранвье ПД распространяется от одного перехвата к другому (скачкообразно) скорость распространения ПД увеличивается.На скорость распространения возбуждения по нервным волокнам влияют следующие факторы:1. Наличие миелиновой оболочки увеличивают скорость.2. Расстояние между перехватами Ранвье - чем он больше, тем больше скорость.Так как в мышечных волокнах отсутствует миелиновая оболочка, то скорость проведения возбуждения по этим волокнам будут определять факторы 3-6.3. Диаметр волокна - чем он больше, тем меньше сопротивление оказывает аксоплазма волокна распространению локальных токов и тем больше скорость проведения возбуждения.4. Амплитуда ПД - чем она больше, тем быстрее деполяризация доходит до Экр, и тем больше скорость проведения.5. Порог деполяризации (ΔV) - чем он меньше, тем быстрее деполяризация мембраны волокна доходит до Экр, и тем больше скорость проведения.6. Скорость нарастания пика ПД - чем она больше, тем быстрее развивается деполяризация к Экр, и тем больше скорость проведения возбуждения.9 ВОПРОСНервно-мышечный синапс образован нервным окончанием аксона -мотонейронов и конечной пластинкой - часть мембраны мышечного волокна, которая контактирует с нервным окончанием. Разделяет пресинаптическую мембрану нервного волкна и конечную пластинку синаптическая щель.М
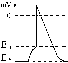
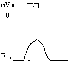 еханизм передачи возбуждениячерез нервно-мышечный синапс состоит в том, что ПД идет по мембране нервного волокна распространяется по пресинаптической мембране, при этом открываются кальциевые каналы пресинаптической мембраны вход ионов Са внутрь нервного окончания взаимодействие с везикулами (пузырьки, в которых медиатор ацетилхолин) движение везикул к пресинаптической мембраны слияния везикул с пресинаптической мембраной выход медиатора ацетилхолина в синаптическую щель диффузия ацетилхолина к мембране конечной пластинки взаимодействие с мембранными циторецепторы (Н-холинорецепторы) открытия хемочутливих натриевых каналов вход ионов Na + в мышечное волокно через конечную пластинку развитие деполяризации концевой пластинки под названием потенциала концевой пластинки (ПКП).
еханизм передачи возбуждениячерез нервно-мышечный синапс состоит в том, что ПД идет по мембране нервного волокна распространяется по пресинаптической мембране, при этом открываются кальциевые каналы пресинаптической мембраны вход ионов Са внутрь нервного окончания взаимодействие с везикулами (пузырьки, в которых медиатор ацетилхолин) движение везикул к пресинаптической мембраны слияния везикул с пресинаптической мембраной выход медиатора ацетилхолина в синаптическую щель диффузия ацетилхолина к мембране конечной пластинки взаимодействие с мембранными циторецепторы (Н-холинорецепторы) открытия хемочутливих натриевых каналов вход ионов Na + в мышечное волокно через конечную пластинку развитие деполяризации концевой пластинки под названием потенциала концевой пластинки (ПКП).
ПКП - один из видов местного возбуждения.Как любое из местных возбуждений ПКП распространяется на соседние участки постсинаптической мембраны с помощью местных токов. Эти токи в невозбуждённых участках мембраны имеют исходное направление, то есть вызывают деполяризацию.Сила этих токов численно соответствует амплитуде ПКП, а она составляет 45-50 мВ. Порог деполяризации постсинаптической мембраны мышечного волокна составляет 40 мВ (величина ПС = -90 мВ, величина Екр = -50 мВ. Следовательно, сила этих токов надпороговая.Время действия этих токов равна продолжительности ПКП и также надпороговая.Скорость увеличения силы соответствует скорости развития ПКП и такжеесть надпороговая.Итак, местные токи в незбуждених участках постсинаптической мембраны вызывают деполяризацию в Екр развитие ПД, которые будут распространяться от места своего возникновения вдоль всей длины мембраны мышечного волокна (и через электромеханическое сопряжение будут вызывать сокращение мышцы). Закономерности проведения возбуждения через нервно-мышечный синапс:1. Одностороннее проведение.2. Замедленное проведения (время от возникновения ПД на пресинаптической мембране к возникновению ПД на постсинаптической мембране составляет около 0,5 мсек.3. Проведение возбуждения через синапс сопровождается быстрым развитием усталости (в связи с истощением запасов медиатора в нервном окончании);Эти особенности (1-3) характерны для любого химического синапса и связанные с химическим характером передачи информации через синапс.4. Возбуждение передается через нервно-мышечный синапс без трансформации ритма - 1 ПД на пресипаптичний мембране вызывает 1 ПКП на мембране концевой пластинки, а это вызывает 1 ПД на мембране мышечного волокна.Блокаторами нервно-мышечной передачи является курареподобные вещества, паприклад, диплацин, образующие устойчивые соединения с Н-холинорецепторами и не позволяют ацетилхолина взаимодействовать с ними. Медиатор ацетилхолин взаимодействует с рецепторами в течение короткого - времени, после чего комплекс медиатор-циторецепторы распадается и медиатор разрушается ферментом ацетилхолинэстеразой, активность которого в синапсе очень высока.10 ВОПРОСВ зависимости от режимов нагрузки выделяют следующие виды мышечного сокращения.1. Изометрического сокращения - сокращение при неизменной довжинни мышцы. Длина мышцы при сокращении остается постоянной, но меняется ее напряжение (Н). В эксперименте такое сокращение можно получить, если изолированный мышцу закрепить с двух сторон и стимулировать электрическим током. В условиях целостного организма изометрическое сокращение бывает,
когда человек пытается, но не может поднять груз.
2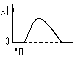 . Изотонический сокращения - сокращение при неизменном напряжении мышцы. Напряжение мышцы при его сокращении остается постоянным, а длина меняется (Δl). В эксперименте такое сокращение можно получить, если один конец изолированного мышцы закрепить, а другой оставить свободным и стимулировать. В условиях целостного организма сокращения мышц максимально приближается к изотонического, если происходит сгибание ненагруженных конечностей.
. Изотонический сокращения - сокращение при неизменном напряжении мышцы. Напряжение мышцы при его сокращении остается постоянным, а длина меняется (Δl). В эксперименте такое сокращение можно получить, если один конец изолированного мышцы закрепить, а другой оставить свободным и стимулировать. В условиях целостного организма сокращения мышц максимально приближается к изотонического, если происходит сгибание ненагруженных конечностей.
3. Смешанное (ауксотоничне) сокращение - сокращение, при котором изменяется длина и напряжение мышцы. Такой тип мышечных сокращений чаще встречается в условиях целостного организма.
В зависимости от частоты стимуляции выделяют следующие виды мышечного сокращения:
1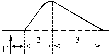 . Одиночное - наблюдается в ответ на единичный стимул или на их серию (если интервал между раздражителями больше или равен длительности одиночного мышечного сокращения). Выделяют следующие периоды мышечного сокращения:
. Одиночное - наблюдается в ответ на единичный стимул или на их серию (если интервал между раздражителями больше или равен длительности одиночного мышечного сокращения). Выделяют следующие периоды мышечного сокращения:
- Латента период (1) - интервал времени от действия раздражителя (П) до начала сокращения;
- Период сокращения (2) - во время этого периода длина мышцы уменьшается;
- Период расслабления (3) - во время этого периода мышца расслабляется, его длина збильшуется к исходному уровню.
2. Тетанических - наблюдается в условиях суммации одиночных мышечных сокращений (одно сокращение накладывается на другое). Такая суммирование возможно, если мышца раздражается серией стимулов и каждый следующий стимул действует тогда, когда предварительное одиночное сокращение еще не закончилось (то есть, интервал между стимулами должен быть меньше продолжительности одиночного мышечного сокращения).
Выделяют следующие виды тетанических сокращений:
Зубчатый тетанус - возникает на базе одиночных сокращений, если каждый следующий стимул попадает на период расслабления одиночого мышечного сокращения. Итак, интервал между стимулами в серии должно быть меньше продолжительности одиночного мышечного сокращения, но больше, чем Сумма продолжительности латентного периода и периода укорочение одиночного сокращения.
Гладкий тетанус - возникает на базе одиночных сокращений, если каждый следующий стимул попадает на латентный период или период укорочение одиночного мышечного сокращения. Итак, интервал между стимулами в серии должен быть меньше или равным сумме продолжительности латентного периода одиночного сокращения, но больше продолжительности латентного периода одиночного сокращения.
Ниже изображен запись регистрации сокращений мышцы при увеличении частоты ее стимуляции. Видно, что при увеличении частоты стимуляции на фазе одиночных сокращений возникают тетанические, сначала зубчатый, а затем гладкой тетанус. Тетанические сокращения отличается от одиночного следующими параметрами:1. Продолжительность тетанического сокращение больше, чем одиночного.2. Амплитуда (сила) тетанического сокращение больше, чем одиночного. Амплитуда гладкого тетануса больше, чем зубчатого.Стрелочками показано момент действия раздражителя на мышечное волокно.Асинхронный тетанус. Асинхронный тетанус представляет вид сокращений, который присущ скелетным мышцам при их естественной деятельности в условиях целостного организма.Каждый скелетная мышца состоит из большого количества отдельных мышечных волокон. При естественной деятельности мышц в условиях целостного организма частота, с которым к ним поступают ПД по аксонам мотонейронов недостаточна для того, чтобы в каждом отдельном волокне мышцы развивался гладкий тетанус. В то же время мышцы человека способны к длительному слитного сокращения. Такое сокращение является следствием асинхронного (неодновременного) сокращение отдельных мышечных волокон мышцы. Каждое волокно сокращается в режиме одиночных сокращений, но в пределах мышцы происходит (добавление) сокращений отдельных волокон. В итоге мышца сокращается слитно и длительно. Это и есть асинхронный тетанус. Его продолжительность может быть очень большой, а его амплитуда не бывает столь значительной, как амплитуда гладкого тетануса, который можно получить в эксперименте.11 ВОПРОСТермин "сопряжение возбуждения с сокращением" означает взаимосвязь возбуждения в скелетных мышцах (возникновение и распространение ПД по мембране волокна) и его сокращение, то есть актомиозиновои взаимодействия.В состоянии покоя взаимодействия актиновых и миозиновых протофибрилл нет потому, что активные центры актина заблокированы регуляторными белками тропонином и тропомиозином. Снимают блокаду ионы Са 2+, концентрация которых в саркоплазме в состоянии покоя низкая 10-8 ммоль / л.