ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 29.10.2023
Просмотров: 159
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
ПД, возникающих в постсинаптической мембране распространяются вдоль всей длины мембраны мышечного волокна, в том числе и по мембране Т-трубочек (это вгинання внешней мембраны мышечных клеток внутрь волокна).
Н
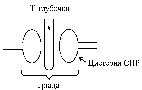 епосредственно рядом с Т-трубочками расположены цистерны саркоплазматического ретикулума (СПР - часть аграпулярнои епдоплазматичнои сетки). Т-трубочки с двумя рядом расположенными цистернами образуют триады. СПР кроме цистерн имеют продольные трубочки, в которых локализуются кальциевые насосы - они активно транспортируют ионы - Са 2+ с саркоплазмы в СПР и в его цистернах накапливается большое количество ионов Са 2+. При движении ПД по мембране Т-трубочек в мембране цистерн СПР открываются кальциевые каналы ионы Са 2+ по градиенту концентрации выходят из цистерн СПР в саркоплазму повышение концентрации ионов Са 2+ в саркоплазме миоцитов с 10-8 до 10-5 ммоль / л диффузия ионов Са 2+ к протофибрилл взаимодействие с регуляторным белком тропонином изменение третичной конформации тропонина и тропомиозином открытия активных центров актина взаимодействие головок миозина с активными центрами актина (мышечное сокращение).
епосредственно рядом с Т-трубочками расположены цистерны саркоплазматического ретикулума (СПР - часть аграпулярнои епдоплазматичнои сетки). Т-трубочки с двумя рядом расположенными цистернами образуют триады. СПР кроме цистерн имеют продольные трубочки, в которых локализуются кальциевые насосы - они активно транспортируют ионы - Са 2+ с саркоплазмы в СПР и в его цистернах накапливается большое количество ионов Са 2+. При движении ПД по мембране Т-трубочек в мембране цистерн СПР открываются кальциевые каналы ионы Са 2+ по градиенту концентрации выходят из цистерн СПР в саркоплазму повышение концентрации ионов Са 2+ в саркоплазме миоцитов с 10-8 до 10-5 ммоль / л диффузия ионов Са 2+ к протофибрилл взаимодействие с регуляторным белком тропонином изменение третичной конформации тропонина и тропомиозином открытия активных центров актина взаимодействие головок миозина с активными центрами актина (мышечное сокращение).В
 основе мышечного сокращения лежит скольжения актиновых протофибрилл относительно миозинових "теория скольжения".
основе мышечного сокращения лежит скольжения актиновых протофибрилл относительно миозинових "теория скольжения".Саркомер - сруктурно функциональный элемент сократительного аппарата скелетных мышц. Они образованы пучками миофибрилл, которые отделены друг от друга перпендикулярными полосами - Z-линиями. До Z-линий прикрепляются одним своим концом тонкие актиновые нити. Другие конце актиновых нитей направлены к центру саркомера и входят в промежутки между толстыми миозиновои нитками. Часть саркомера, примыкающая к Z линии и образована только актиновыми протофибрилл, называется I-дисков (изотропных) вслед за ними расположены А-диски (анизотропные) - часть саркомера, где имеет место взаимное перекрытие актиновых и миозиновых протофибрилл. При укорочении мышцы, в, ходе его сокращение, укорачивается длина протофибрилл укорочение длины каждого саркомера. Но при этом длина анизотропных дисков не уменьшается, а уменьшается длина изотропных дисков. Это является следствием скольжения актиновых протофибрилл относительно миозинових по направлению к центру саркомера.
П
 ричиной движения актиновых нитей относительно миозинових (их скольжения) является "наклон" головки после ее присоединения к активному центру актина. Большое количество головок, входящих в состав актомиозинових мостиков, одновременно делают "наклон" нить актина перемещается к центру саркомера вдоль нити миозина. За счет одного такого "наклона" саркомер (мышца) может укоротиться на 1% своей исходной длины. Степень укорочение мышцы может достигать 50% такие "наклонности" головок во время одного мышечного сокращения должны повториться 50 раз. Это возможно, если актомиозин мостики после "наклона" распадаются (диссоциируют) головка миозина взаимодействует с последующим активным центром актина следующий "наклон" распад мостике и т.д. На "наклон" одной головки миозина расходуется энергия одной молекулы АТФ. Для того, чтобы произошел распад актомиозинового мостика необходимо, чтобы к головке миозина присоединилась молекула АТФ (но энергия АТФ при этом не расходуется).
ричиной движения актиновых нитей относительно миозинових (их скольжения) является "наклон" головки после ее присоединения к активному центру актина. Большое количество головок, входящих в состав актомиозинових мостиков, одновременно делают "наклон" нить актина перемещается к центру саркомера вдоль нити миозина. За счет одного такого "наклона" саркомер (мышца) может укоротиться на 1% своей исходной длины. Степень укорочение мышцы может достигать 50% такие "наклонности" головок во время одного мышечного сокращения должны повториться 50 раз. Это возможно, если актомиозин мостики после "наклона" распадаются (диссоциируют) головка миозина взаимодействует с последующим активным центром актина следующий "наклон" распад мостике и т.д. На "наклон" одной головки миозина расходуется энергия одной молекулы АТФ. Для того, чтобы произошел распад актомиозинового мостика необходимо, чтобы к головке миозина присоединилась молекула АТФ (но энергия АТФ при этом не расходуется).
Для расслабления скелетной мышцы необходимо, чтобы уменьшилась концентрация ионов Са 2+ в саркоплазме. Это достигается благодаря действию кальциевых насосов продольных трубочек СПР активный транспорт ионов Са 2+ в СПР уменьшение его концентрации в саркоплазме до уровня 10-8 ммоль / л отсоединения ионов Са 2+ от регуляторного белка тропонина восстановления исходной третичной структуры тропонина и тропомиозином блокада активных центров актина расслабление мышцы.
Таким образом, во время мышечного сокращения энергия АТФ расходуется:
1. На работу натрий-калиевого насоса внешней мембраны мышечного волокна (этот насос поддерживает градиент концентрации ионов К + и Na + вне и внутри клетки, что необходимо для поддержания на необходимом уровне ПС и для развития ПД на мембране).
2. На "наклон" головки миозина.
3. На работу кальциевого насоса СПР расслабление мышцы.
12 ВОПРОС
Гладкие мышцы находятся в стенках внутренних органов и кровеносных сосудов. Регуляция их тонуса и сократительной активности осуществляется эфферентными волокнами симпатической и парасимпатической нервной системы, а также местными гуморальными и физическими воздействиями.
Сократительный аппарат гладких мышц, как и скелетных, состоит из толстых миозиновых и тонких актиновых нитей. Вследствие их нерегулярного распределения клетки гладких мышц не имеют характерной для скелетной и сердечной мышцы поперечной исчер-ченности. Гладкомышечные клетки имеют веретенообразную форму, длину 50-400 мкм и толщину 2-10 мкм. Они отделены друг от друга узкими щелями (60-150 нм). Возбуждение электротонически распространяется по мышце от клетки к клетке через особые плотные контакты (нексусы)между плазматическими мембранами соседних клеток.
Волокна гладких мышц сокращаются в результате относительного скольжения миозиновых и актиновых нитей, но скорость их сокращения и скорость расщепления АТФ в 100-1000 раз меньше, чем в скелетных мышцах. Поэтому гладкие мышцы хорошо приспособлены к длительному тоническому сокращению без развития утомления. При этом их энерготраты крайне невелики. По своим функциональным особенностям гладкие мышцы подразделяются на мышцы, обладающие и не обладающие спонтанной активностью.
Гладкие мышцы, обладающие спонтанной активностью
Гладкие мышцы, обладающие спонтанной активностью, способны сокращаться и при отсутствии прямых возбуждающих нервных и гуморальных воздействий (например, ритмические сокращения гладких мышц кишечника). Потенциал покоя таких клеток постоянно спонтанно колеблется в пределах 30-70 мВ («дрейф»). В том случае, когда потенциал покоя в результате такого «дрейфа» снижается до некоего критического уровня, возникает потенциал действия, вызывающий сокращение мышечного волокна. Длительность потенциала действия гладкомышечных волокон намного больше, чем у волокон поперчено-полосатой мускулатуры (несколько секунд). Продолжительность самого сокращения также превышает несколько секунд. Особенно медленно протекает расслабление. Возникающее в одной группе клеток возбуждение распространяется по всей мышце со скоростью не превышающей 2- 10 см/с, то есть намного меньшей, чем по скелетной мышце.
Спонтанная активность гладкомышечных клеток связана и с их растяжением, вызывающим деполяризацию мембраны мышечного волокна, возникновение серии распространяющихся потенциалов действия, с последующим сокращением клетки.
Гладкие мышцы, не обладающие спонтанной активностью
Гладкие мышцы, не обладающие спонтанной активностью сокращаются под влиянием импульсов вегетативной нервной системы. Так, в отличие от мышц кишечника, мышечные клетки артерий, семенных протоков и радужки обладают слабой спонтанной активностью, илл вообще не проявляют ее. Отдельные нервные импульсы не способны вызвать пороговую деполяризацию таких клеток и их сокращение. Потенциал действия волокна с последующим сокращением возникает лишь при поступлении к нему серии импульсов с частотой 1 имп/с и выше. В гладких мышцах, не обладающих спонтанной активностью возбуждение также передается от одной клетки к последующим через плотные контакты их мембран.
13 ВОПРОС
Утомление мышц. Утомление - временное снижение или потеря работоспособности отдельной клетки, ткани, органа или организма в целом, наступающее после нагрузок (деятельности). Утомление мышц происходит при их длительном сокращении (работе) и имеет определенное биологическое значение, сигнализируя о истощении (частичном) энергетических ресурсов.
При утомлении понижаются функциональные свойства мышцы: возбудимость, лабильность и сократимость. Высота сокращения мышцы при развитии утомления постепенно снижается. Это снижение может дойти до полного исчезновения сокращений. Понижаясь, сокращения делаются все более растянутыми, особенно за счет удлинения периода расслабления: по окончании сокращения мышца долго не возвращается к первоначальной длине, находясь в состоянии контрактуры (крайне замедленное расслабление мышцы). Скелетные мышцы утомляются раньше гладких. В скелетных мышцах сначала утомляются белые волокна, а потом красные.
Теория истощения,предложенная К. Шиффом. Согласно этой теории причиной утомления служит исчезновение в мышце энергетических веществ, в частности гликогена.
Теория засорения органа продуктами рабочего распада (теория отравления). Согласно этой теории, утомление объясняется накоплением большого количества молочной, фосфорной кислот и недостатком кислорода, а так же других продуктов обмена, которые нарушают обмен веществ в работающем органе и его деятельность прекращается.
в утомленной мышце появляются продукты обмена веществ, уменьшается содержание гликогена, АТФ, креатинофосфата. Изменения наступают в сократительных белках мышцы. Происходит связывание или уменьшение сульфгидрильных групп актомиозина, в результате чего нарушается процесс синтеза и распада АТФ.
Быстрая утомляемость синапсов обусловлена несколькими факторами.
Во-первых, при длительном раздражении в нервных окончаниях уменьшается запас медиатора, а его синтез не поспевает за расходованием.
Во-вторых, накапливающиеся продукты обмена в мышце понижают чувствительность постсинаптической мембраны к ацетилхолину, в результате чего уменьшается величина постсинаптического потенциала. Когда он понижается до критического уровня, в мышечном волокне не возникает возбуждения.
14 ВОПРОС
Нейрон — это нервная клетка с отростками, являющаяся основной структурной и функциональной единицей нервной системы. Она имеет строение, сходное с другими клетками: оболочка, протоплазма, ядро, митохондрии, рибосомы и другие органоиды.
Основные свойства нейронов
: раздражимость, возбудимость, проводимость, лабильность, инертность, утомляемость, торможение, регенерация и др.
Раздражимость — способность нервной клетки отвечать на различные раздражения биохимическими изменениями, сопровождающимися нарушением ионного равновесия и деполяризацией электрических зарядов на мембранах клетки в месте раздражения.
Возбудимость — способность отдельных частей нервной клетки генерировать электрохимические импульсы, т. е. отвечать на раздражение возбуждением.
Проводимость — способность нейрона проводить импульсы возбуждения с определенной скоростью, в неизменном ритме и силе.
Лабильность (подвижность) — способность нервной клетки принимать и передавать максимальное число импульсов за единицу времени без искажения.
В результате длительной эволюции нервные клетки стали выполнять следующие основные функции: рецепторную, сенсорную, информационную и моторную.
15 ВОПРОС
Біологічна регуляція
Внутрішньоклітинна
Організмова
НЕРВОВА
ГУМОРАЛЬНА
ГЕННА
ФЕРМЕНТАТИВНА
МІОГЕННА
Биологическая регуляция - это совокупность процессов, обеспечивающих взаимодействие структур организма и взаимодействие организма с внешней средой при достижении его приспособительной реакции.
Контур регуляции - это путь передачи и обработки информации, обеспечивающий достижение приспособительной реакции организма.
По способам передачи информации различают два вида регуляции: а) нервная регуляция, б) гуморальная регуляция: