ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.01.2024
Просмотров: 103
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
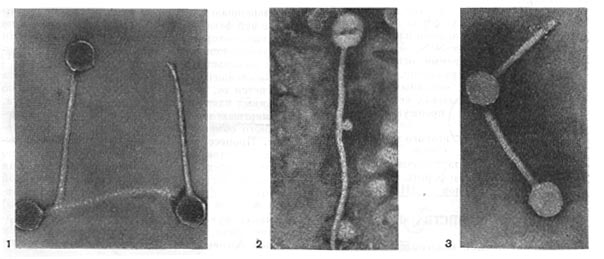
Рис.6. Разные фаги пятого морфологического типа, частица состоит из головки и длинного отростка чехол которого не способен сокращаться. 1,2- увел. X 225 000, 3 - увел. X250 000
Нуклеиновая кислота находится в головке. Внутри головки фагов обнаружено также небольшое количество белка (около 3%).
Таким образом, по химическому составу фаги являются нуклеопротеидами. В зависимости от типа своей нуклеиновой кислоты фаги делятся на ДНК-овые и РНК-овые. Количество белка и нуклеиновой кислоты у разных фагов разное. У некоторых фагов содержание их почти одинаковое и каждый из этих компонентов составляет около 50%. У других фагов соотношение между этими основными компонентами может быть различно.
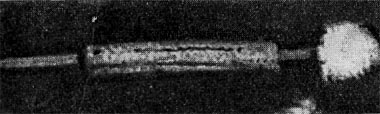
Рис.7. Фаг шестого морфологического типа, частица состоит из головки и длинного отростка, чехол которого способен к сокращению. Увел, около 400 000.
Кроме указанных основных компонентов, фаги содержат в небольших количествах углеводы и некоторые преимущественно нейтральные жиры.
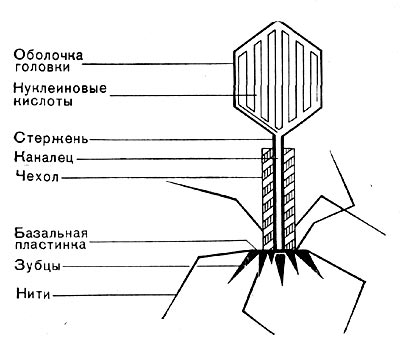
Рис.8. Схема строения фаговой частицы.
Все известные фаги второго морфологического типа РНК-овые. Среди фагов третьего морфологического типа встречаются как РНК-овые, так и ДНК-овые формы. Фаги остальных морфологических типов - ДНК-овые.
6 Жизненный цикл
Электронный микроскоп открыл тайну воспроизведения вирусов. Проще всего этот процесс наблюдать на группе вирусов, нападающих на бактерии, - бактериофагов, или просто фагов. Фаги легко размножаются в кишечной палочке Escherichiacoli. Ее клетки поражаются по меньшей мере семью штаммами фаговТ1-Т7. Природа позаботилась об этих вирусах, снабдив их необходимыми орудиями: ферментом лизоцимом, «открывателем» клеточной мембраны, и микроскопическими «мышцами», сокращение которых способствует попаданию ДНК внутрь клетки.
Электронная микрофотография бактериофагов, инфицирующих клетку
Атака начинается с прикрепления хвостов нескольких десятков фагов к определенной части бактериальной стенки. Тотчас же лизоцим растворяет клеточную мембрану. Аппарат хвоста вируса действует наподобие шприца: «мышцы» сокращаются и нуклеиновая кислота впрыскивается внутрь клетки. Верхняя «одежда» вируса - белковый чехол - остается снаружи клетки. Так завершается «оккупация» бактерий фагами. Весь процесс длится всего несколько минут. Нуклеиновая кислота играет главную роль в воспроизведении фага. Это было доказано немецкими учеными в опытах по гибридизации вирусов. Однако не все вирусы так агрессивны, как фаги, не у всех есть хвост с набором необходимых для взлома клетки инструментов.
Как же в этих случаях вирус проникает на чужую территорию? Как ни парадоксально, в этих случаях сама клетка осуществляет внедрение вирусов - она как бы заглатывает их. На протяжении эволюции у клеток выработался механизм активного захвата из окружающей среды различных твердых частиц (фагоцитоз) и капелек жидкости (пиноцитоз). Этот механизм играет большую роль в нормальной жизнедеятельности клетки. В случае же встречи с вирусом происходит как бы «самоубийство» клетки.
Участие обратной транскриптазы в образовании комплиментарной ДНК на вирусной одноцепочечной РНК-матрице в животной клетке
При «заглатывании» вируса клетка стремится «переварить» его специальными ферментами клетки. «Война» может быть закончена миром, если клетке не удастся подобрать ключ - соответствующий фермент для растворения белковой оболочки. В случае же наличия в клетке подходящего фермента происходит следующее: фермент разрушит оболочку вируса, освободит нуклеиновую кислоту, которая по клеточным каналам быстро доберется до ядра клетки и проникнет в него или останется в цитоплазме. Вот с этого момента начинается размножение вируса. Далее клетка будет разрушена. Далее вирусная нуклеиновая кислота, подавляя производство молекул клетки, вынуждает клетку производить вирусы. Вначале оккупированная клетка начинает синтезировать ферменты - полимеразы, снимающие копии с нуклеиновой кислоты вируса. Таким образом происходит редупликация, совершающаяся в разных частях клетки, в зависимости от вируса. При образовании достаточного количества вирусных нуклеиновых кислот часть из них отправляется на производство белков клетки. Рибосомы теперь вынуждены синтезировать вирусные белки вместо собственных. Оставшаяся часть вирусных нуклеиновых кислот идет на производство дочерних вирусов.
7 Размножение фагов
Взаимоотношения между фагом и чувствительной к нему клеткой очень сложны и не всегда завершаются лизисом клетки и размножением в ней фага. Мы рассмотрим такую инфекцию клетки, которая заканчивается гибелью клетки и размножением в ней фага. Такая инфекция называется продуктивной.
Важнейшей особенностью размножения фага является то, что оно может происходить только в живых клетках, находящихся в стадии роста. В мертвых клетках, а также продуктах клеточного обмена размножение фага не происходит. Процесс размножения фага весьма сложный и состоит из следующих последовательно протекающих этапов (рис. 9):
1) адсорбция фаговой частицы на поверхности микробной клетки;
2) проникновение содержимого головки фаговой частицы (нуклеиновой кислоты) в микробную клетку;
3) внутриклеточное развитие фага, заканчивающееся образованием новых фаговых частиц;
4) лизис клетки и выход из нее новых фагов.
Время с момента инфицирования клетки фагом до лизиса клетки называется латентным или скрытым периодом. Продолжительность этого периода различна для разных типов фага, зависит от окружающей температуры, состава среды и других факторов. Латентный период фагов, специфичных для одних бактерий, 15-40 мин, для других - 5 ч и более. У фагов актиномицетов латентный период может быть еще продолжительнее. При низкой температуре латентный период значительно увеличивается.
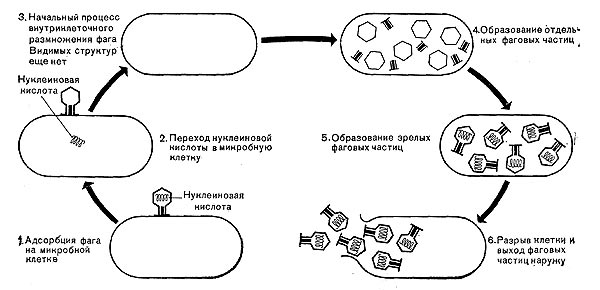
Рис.9. Схема размножения фага.
Из всех этапов размножения фага наиболее изучен первый - адсорбция.
Адсорбция фага на клетке - реакция весьма специфичная. В клеточной стенке бактерий имеются особые структуры (рецепторы), к которым могут прикрепиться фаги. Адсорбируются на рецепторах только те фаги, к которым чувствительна клетка. Фаги, имеющие отростки, прикрепляются к микробной стенке свободным концом отростка. Нитевидные фаги, а также фаги, не имеющие отростков, адсорбируются не на микробной стенке, а на нитевидных структурах, окружающих стенку, - фимбриях. Описаны фаги, которые прикрепляются отростком к бактериальным жгутикам. У некоторых фагов процесс адсорбции может осуществляться лишь в том случае, когда в среде имеются определенные вещества - кофакторы: аминокислоты (триптофан, тирозин и др.) или соли (кальциевые, магниевые).
На конце фагового отростка имеется особый фермент типа лизоцима. После адсорбции фага под влиянием этого фермента происходит растворение стенки микробной клетки и содержимое головки фага - нуклеиновая кислота - перекачивается в микробную клетку. Этим завершается второй этап процесса размножения фага.
Остальные структуры фаговой частицы - оболочка головки, отросток и его субструктуры - внутрь инфицированной фагом клетки не попадают. Их роль заключается в обеспечении сохранности фаговой частицы, находящейся вне клетки, и содействии проникновению фаговой нуклеиновой кислоты в клетку при инфекции.
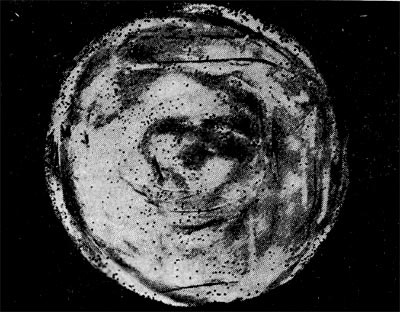
Рис.10. Мелкие (едва видимые) негативные колонии актинофага. Увел. 6 : 10.
У нитевидных фагов, в отличие от других видов фагов, внутрь клетки проникает весь белок или его часть. После проникновения нуклеиновой кислоты фага в клетку начинается сложный процесс внутриклеточного размножения фага. Под влиянием нуклеиновой кислоты фага резко изменяется весь обмен микробной клетки. Основные процессы, протекающие в инфицированной клетке, направлены на образование новых фаговых частиц. Вначале формируются отдельно головки и отростки, которые затем объединяются в зрелые фаговые частицы. К этому времени внутри клетки образуется особый литический фермент, который вызывает лизис клетки изнутри. Клетка распадается, и новые зрелые частицы фага выходят наружу
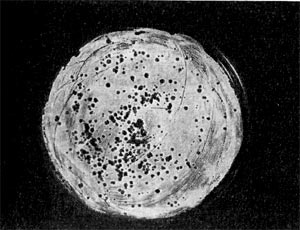
Рис. 11 Мелкие негативные колонии актинофага Увел. 8 : 9.
Количество новых фаговых частиц, образуемых одной клеткой при фаговой инфекции, называют выходом фага или его урожайностью. Выход фага зависит от свойств данного фага и не зависит от клетки-хозяина и ее размеров. Одни фаги отличаются очень низким выходом (5-50 частиц на клетку), у других выход значительно выше (от 1000 до 2500). Особенно высоким выходом отличаются мелкие РНК-овые фаги (свыше 20 000 частиц на клетку). Если большое количество бактериальных клеток смешать с небольшим количеством фаговых частиц, то процесс размножения фагов проходит несколько циклов. Вначале инфицируется часть клеток. Первое потомство фага инфицирует оставшиеся клетки - происходит второй цикл, за ним может следовать третий и т. д., пока не будут лизированы все чувствительные к данному фагу клетки. Среди фагов встречаются такие, размножение которых возможно лишь при наличии в среде определенных кофакторов. Одни из этих веществ, как уже указывалось, необходимы для адсорбции фага; другие - для внутриклеточного размножения фага.
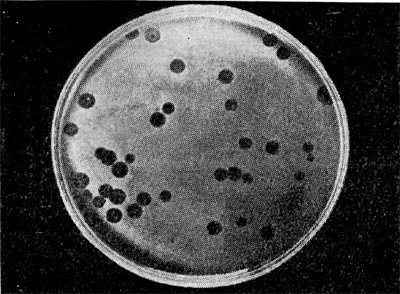
Рис.12. Негативные колонии бактериофага средней величины. Увел. 1:1.
Большинство фагов вызывают при размножении лизис клетки и ее гибель. Лишь в последнее время было установлено, что при инфицировании клетки нитевидными фагами возможно размножение их без гибели клетки.
Если произвести рассев по поверхности агаризованной питательной среды в чашках Петри смеси фага и чувствительных к нему микробов и чашки выдержать в термостате, то происходит лизис клеток в результате размножения фага. Если взять большое количество частиц фага, то лизируется большая часть или весь выросший газон культуры. Если количество фаговых частиц таково, что они распределяются только на отдельных участках газона, лизируя в этих местах культуру, то возникает колония фага.
Эти колонии фага получили название бляшек, стерильных пятен. Правильнее их называть негативными колониями. Каждая негативная колония состоит из десятков и сотен миллионов фаговых частиц. Размер негативных колоний и их форма зависят в первую очередь от свойств фага, а также от состава среды и культуры микробов. У одних фагов негативные колонии очень мелкие и еле видимы невооруженным глазом, другие достигают 10 мм в диаметре и более. Колонии бывают светлые и четкие, когда лизировалась вся культура, или мутноватые, когда лизировались не все клетки. Вокруг негативных колоний некоторых фагов могут возникнуть различной формы и величины ореолы. На рисунках 10-14 показаны негативные колонии разных фагов. Морфология негативных колоний служит одним из признаков, которым пользуются при дифференциации фагов.
8 Распространение фагов
В настоящее время найдены, фаги, лизирующие клетки микроорганизмов, принадлежащих ко всем систематическим группам, как патогенных для человека, животных и растений, так и сапрофитных (непатогенных).
До недавнего времени не было ясно, существуют ли фаги против плесневых грибов и дрожжей. В последние годы найдены фаги, активные против грибов родов пенициллов, аспергиллов и других, а также против некоторых дрожжей. Интересно отметить, что вирус удалось выявить и у тех видов пенициллов, которые применяются в промышленности для получения пенициллина. Не выявлены вирусы, активные против простейших животных, а также истинных спирохет.
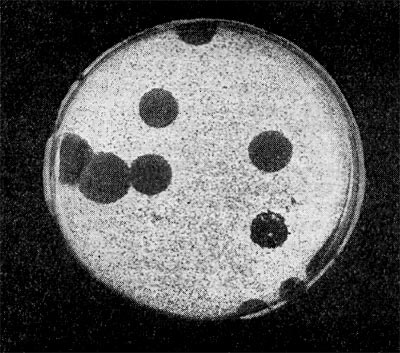
Рис.13. Крупные негативные колонии актинофага. Увел. 1:1.
В природных условиях фаги встречаются в тех местах, где есть чувствительные к ним бактерии. Чем богаче тот или иной субстрат (почва, вода, выделения человека и животных и т. д.) микроорганизмами, тем в большем количестве в нем встречаются соответствующие фаги. Так, фаги, лизирующие клетки всех видов почвенных микроорганизмов, находятся в почвах. Особенно богаты фагами черноземы и почвы, в которые вносились органические удобрения. Фаги, активные против разных видов кишечной, дизентерийной, тифозной и паратифозной палочек, часто встречаются в содержимом кишечника человека и животных, сточных водах и загрязненных водоемах. Фаги фитопатогенных микроорганизмов успешнее всего выделяются из остатков растений, пораженных этими микробами.