Файл: Учебник для высших учебных заведений физической культуры Издание 2е, исправленное и дополненное.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.01.2024
Просмотров: 2067
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
новых мостиков на следующем участке актиновой нити. В результате повторения подобных процессов многократного образования и распада мостиков сокращается длина отдельных саркомеров и всего мышечного волокна в целом. Максимальная концентрация кальция в миофибрилле достигается уже через 3 мс после появления потенциала действия в поперечных трубочках, а максимальное напряжение мышечного волокна — через 20 мс.
Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью (или электромеханическим сопряжением). В результате сокращения мышечного волокна актин и миозин более равномерно распределяются внутри саркомера, и исчезает видимая под микроскопом поперечная исчерченность мышцы.
Расслабление мышечного волокна связано с работой особого механизма — «кальциевого насоса», который обеспечивает откачку ионов Са из миофибрилл обратно в трубочки саркоплазматического ретикулума. На это также тратится энергия АТФ.
из миофибрилл обратно в трубочки саркоплазматического ретикулума. На это также тратится энергия АТФ.
5.3. ОДИНОЧНОЕ И ТЕХНИЧЕСКОЕ СОКРАЩЕНИЕ. ЭЛЕКТРОМИОГРАММА
При единичном надпороговом раздражении двигательного нерва или самой мышцы возбуждение мышечного волокна сопровождается
одиночным сокращением. Эта форма механической реакции состоит из 3 фаз: латентного или скрытого периода, фазы сокращения и фазы расслабления. Самой короткой фазой является скрытый период, когда в мышце происходит электромеханическая передача. Фаза расслабления обычно в 1.5-2 раза более продолжительна, чем фаза сокращения, а при утомлении затягивается на значительное время.
Если интервалы между нервными импульсами короче, чем длительность одиночного сокращения, то возникает явление суперпозиции — наложение механических эффектов мышечного волокна друг на друга и наблюдается сложная форма сокращения — тетанус. Различают 2 формы тетануса — зубчатый тетанус, возникающий при более редком раздражений, когда происходит попадание каждого следующего нервного импульса в фазу расслабления отдельных оди ночных сокращений, и сплошной или гладкий тетанус, возникающий при более частом раздражении, когда каждый следующий импульс попадает в фазу сокращения (рис. 11). Таким образом, (в некоторых границах) между частотой импульсов возбуждения и амплитудой сокращения волокон ДЕ существует определенное соотношение: при небольшой частоте (например, 5-8 имп. в 1с)
54
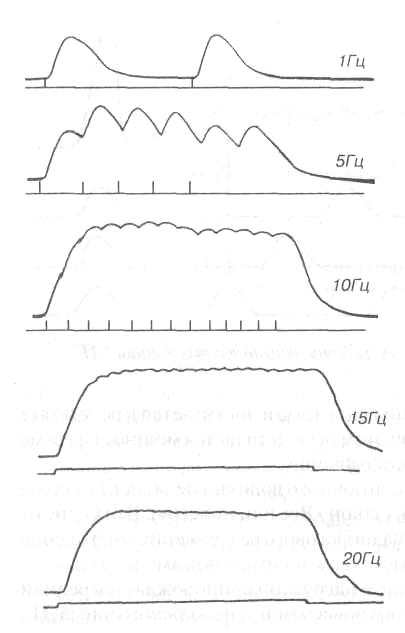
Рис. П. Одиночноесокращение, зубчатыйи сплошной тетанускамбаловидпой мышцычеловека(по: Зимкин Н.В. и др., 1984). Верхняя кривая — сокращение мышцы, нижняя — отметкараздражения мышцы,справа указана частотараздражения
возникают одиночные сокращения, при увеличении частоты (15-20 имп. в 1с) — зубчатый тетанус, при дальнейшем нарастании частоты (25-60 имп. в 1 с) — гладкий тетанус. Одиночное сокращение — более слабое и менее утомительное, чем тетаническое. Зато тетанус обеспечивает в несколько раз более мощное, хотя и кратковременное сокращение мышечного волокна.
Сокращение целой мышцы зависит от формы сокращения отдельных ДЕ и их координации во времени. При обеспечении длительной, но не очень интенсивной работы, отдельные ДЕ сокращаются попеременно (рис. 12), поддерживая общее напряжение мышцы на заданном уровне (например, при беге на длинные и сверхдлинные дистанции). При этом отдельные ДЕ могут развивать как одиночные, так и тетанические сокращения, что зависит от частоты нервных импульсов. Утомление в этом случае развивается медленно, так как, работая по очереди, ДЕ в промежутках между активацией успевают восстанавливаться. Однако для мощного кратковременного усилия (например, поднятия штанги) требуется синхронизация активности отдельных ДЕ, т. е. одновременное возбуждение практически всех ДЕ. Это, в свою очередь, требует одновременной активации
55
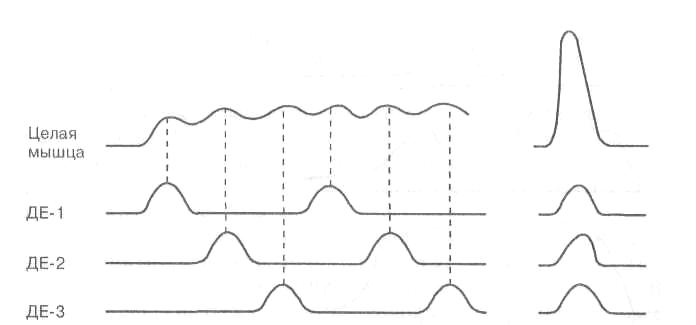
Рис. 12. Различные режимы работы двигательных единиц (ДЕ)
соответствующих нервных центров и достигается в результате длительной тренировки. При этом осуществляется мощное и весьма утомительное тетаническое сокращение.
Амплитуда сокращения одиночного волокна не зависит от силы надпорогового раздражения (закон «Все или ничего»). В отличие от этого, при нарастании силы надпорогового раздражения сокращение целой мышцы постепенно растет до максимальной амплитуды.
Работа мышцы с небольшой нагрузкой сопровождается редкой частотой нервных импульсов и вовлечением небольшого числа ДЕ. В этих условиях, накладывая отводящие электроды на кожу над мышцей и используя усилительную аппаратуру, можно на экране осциллографа или с применением чернильной записи на бумаге зарегистрировать одиночные потенциалы действия отдельных Д Е. В случае же значительных напряжений потенциалы действия многих ДЕ алгебраически суммируются и возникает сложная интегрированная кривая записи электрической активности целой мышцы — электромиограмма (ЭМГ).
Форма ЭМГ отражает характер работы мышцы: при статических усилиях она имеет непрерывный вид, а при динамической работе — вид отдельных пачек импульсов, приуроченных, в основном, к начальному моменту сокращения мышцы и разделенных периодами «электрического молчания». Особенно хорошо ритмичность появления подобных пачек наблюдается у спортсменов при циклической работе (рис. 13). У маленьких детей и неадаптированных к такой работе лиц четких периодов отдыха не наблюдается, что отражает недостаточное расслабление мышечных волокон работающей мышцы.
Чем больше внешняя нагрузка и cилa сокращения мышцы, тем выше амплитуда ее ЭМГ. Это связано с увеличением частоты нервных импульсов, вовлечением большего числа ДЕ в мышце и синхронизацией
56
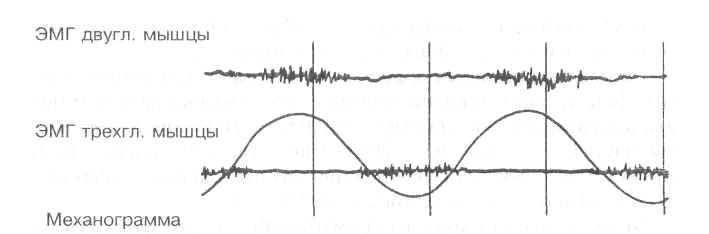
Рис. 13. Электромиограмма мышц-антагонистов при циклической работе
их активности. Современная многоканальная аппаратура позволяет производить одновременную регистрацию ЭМГ многих мышц на разных каналах. При выполнении спортсменом сложных движений можно видеть на полученных ЭМГ кривых не только характер активности отдельных мышц, но и оценить моменты и порядок их включения или выключения в различные фазы двигательных актов. Записи ЭМГ, полученные в естественных условиях двигательной деятельности, можно передавать к регистрирующей аппаратуре по телефону или радиотелеметрически. Анализ частоты, амплитуды и формы ЭМГ (например, с помощью специальных компьютерных программ) позволяет получить важную информацию об особенностях техники выполняемого спортивного упражнения и степени ее освоения обследуемым спортсменом.
По мере развития утомления при той же величине мышечного усилия амплитуда ЭМГ нарастает. Это связано с тем, что снижение сократительной способности утомленных ДЕ компенсируется нервными центрами вовлечением в работу дополнительных ДЕ, т. е. путем увеличения количества активных мышечных волокон. Кроме того, усиливается синхронизация активности ДЕ, что также повышает амплитуду суммарной ЭМГ.
Движение является результатом взаимодействия внутренних и внешних сил, развиваемых в опорно-двигательном аппарате, — активных (возникающих при сокращении или напряжении мышцы во время ее возбуждения) и пассивных (упругое напряжение при растяжении мышцы, сопротивление мышцы и ее сухожилия).
Сила мышцы зависит отряда морфологических и физиологических факторов: количества и свойств мышечных волокон в мышце,
57
исходной длины мышцы, характера нервных импульсов, механических условий действия мышцы на кости скелета.
Сила мышцы является суммой силы отдельных ее мышечных волокон. Подсчитано, что 1 одиночное мышечное волокно икроножной мышцы развивает напряжение 100-200 мг, ІДЕ икроножной мышцы человека содержит около 2000 мышечных волокон и развивает напряжение 200-400 г, 1 икроножная мышца содержит около 1000 ДЕ, т. е. развивает напряжение 200-400 кг.
Большое значение имеет анатомическое строение мышцы. В параллельно-волокнистых и веретенообразных мышцах (камбаловидная мышца и др.) сила мышц тем больше, чем больше ее анатомический поперечник, т. е. площадь поперечного сечения целой мышцы. В перистых мышцах (двуглавая мышца и др.) физиологический поперечник, т. е. площадь поперечного сечения всех мышечных волокон гораздо больше, чем ее анатомический поперечник. В такой мышце упаковано значительно больше мышечных волокон и, соответственно, больше ее сила.
На силу сокращения мышцы влияет ее исходная длина, так как от нее. зависит возможное количество поперечных мостиков между актином и миозином. Предполагают, что в каждом цикле присоединения-отсоединения поперечных мостиков расходуется энергия 1 молекулы АТФ на 1 поперечный мостик. Следовательно, чем больше образуется в мышечном волокне актино-миозиновых мостиков, тем выше скорость расщепления АТФ, больше тяга сократительных белков и, соответственно, больше развиваемая мышцей сила.
Наибольшее количество актино-миозиновых контактов образуется при небольшом растяжении мышцы до некоторой оптимальной длины. При значительном растяжении саркомера нити актина далеко расходятся в стороны и практически не контактируют с расположенным в средней части саркомера миозином. В случае же резкого уменьшения длины саркомера нити актина в центре перекрывают друг друга, препятствуя контактам с миозином и также уменьшая число образуемых мостиков. В связи с этими особенностями взаимодействия сократительныхбелков наибольшая сила мышцы проявляется при некотором ее предварительном растяжении.
Одной из важнейших характеристик скелетных мышц, влияющих на силу сокращения, является состав (композиция) мышечных волокон. Различают 3 типа мышечных волокон — медленные неутомляемые (I типа), быстрые неутомляемые или промежуточные ( 11-а типа) и быстрые утомляемые (11-б типа).
Медленные волокна (1 типа), их обозначают также SO — Slow Oxydative (англ. — медленные окислительные) — это выносливые (неутомляемые) и легко возбудимые волокна, с богатым кровоснабжением, большим количеством митохондрий, запасов миоглобина и
58
с использованием окислительных процессов энергообразования (аэробные). Их, в среднем, у человека 50%. Они легко включаются в работу при малейших напряжениях мышц, очень выносливы, но не обладают достаточной силой. Чаще всего они используются при поддержании ненагрузочной статической работы, например, при сохранении позы.
Быстрые утомляемые волокна (11-б типа) или FG — Fast Glicolitic (быстрые гликолитические) используют анаэробные процессы энергообразования (гликолиз). Они менее возбудимы, включаются при больших нагрузках и обеспечивают быстрые и мощные сокращения мышц. Зато эти волокна быстро утомляются. Их примерно 30%. Волокна промежуточного типа (П-а) — быстрые неутомляемые, окислительные, их около 20%. В среднем, для разных мышц характерно различное соотношение медленных неутомляемых и быстрых утомляемых волокон. Так, в трехглавой мышце плеча преобладают быстрые волокна (67%) над медленными (33%), что обеспечивает скоростно-силовые возможности этой мышцы (рис. 14), а для более медленной и выносливой камбаловидной мышцы характерно наличие 84% медленных и всего 16% быстрых волокон (Салтан Б., 1979).
Однако, состав мышечных волокон в одной и той же мышце имеет огромные индивидуальные различия, зависящие от врожденных типологических особенностей человека. К моменту рождения человека его мышцы содержат лишь медленные волокна, но под влиянием нервной регуляции устанавливается в ходе онтогенеза генетически заданное индивидуальное соотношение мышечных волокон разного типа. По мере перехода от зрелого возраста к пожилому число быстрых волокон у человека заметно снижается и, соответственно, уменьшается мышечная сила. Например, наибольшее количество быстрых волокон в наружной головке 4-х главой мышцы бедра мужчины (около 59-63%) отмечается в возрасте 20-40 лет, а в возрасте 60-65 лет их число почти на 1/3 меньше (45%).
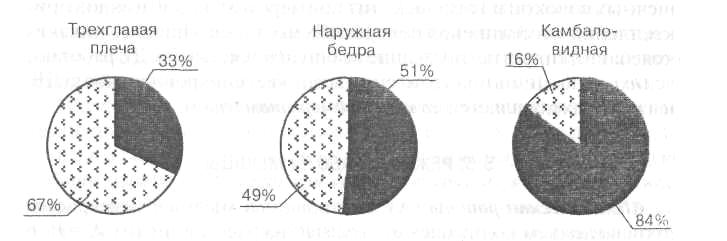
Рис. 14. Состав мышечных волокон в разных мышцах
Медленные — черным цветом; быстрые — серым
59
Количество тех или других мышечных волокон не изменяется в процессе тренировки. Возможно только нарастание толщины (гипертрофия) отдельных волокон, а также некоторое изменение свойств промежуточных волокон. При направленности тренировочного процесса на развитие силы происходит нарастание объема быстрых волокон, что и обеспечивает повышение силы тренируемых мышц.
Весь процесс от появления мышечного потенциала действия до сокращения мышечного волокна называется электромеханической связью (или электромеханическим сопряжением). В результате сокращения мышечного волокна актин и миозин более равномерно распределяются внутри саркомера, и исчезает видимая под микроскопом поперечная исчерченность мышцы.
Расслабление мышечного волокна связано с работой особого механизма — «кальциевого насоса», который обеспечивает откачку ионов Са
5.3. ОДИНОЧНОЕ И ТЕХНИЧЕСКОЕ СОКРАЩЕНИЕ. ЭЛЕКТРОМИОГРАММА
При единичном надпороговом раздражении двигательного нерва или самой мышцы возбуждение мышечного волокна сопровождается
одиночным сокращением. Эта форма механической реакции состоит из 3 фаз: латентного или скрытого периода, фазы сокращения и фазы расслабления. Самой короткой фазой является скрытый период, когда в мышце происходит электромеханическая передача. Фаза расслабления обычно в 1.5-2 раза более продолжительна, чем фаза сокращения, а при утомлении затягивается на значительное время.
Если интервалы между нервными импульсами короче, чем длительность одиночного сокращения, то возникает явление суперпозиции — наложение механических эффектов мышечного волокна друг на друга и наблюдается сложная форма сокращения — тетанус. Различают 2 формы тетануса — зубчатый тетанус, возникающий при более редком раздражений, когда происходит попадание каждого следующего нервного импульса в фазу расслабления отдельных оди ночных сокращений, и сплошной или гладкий тетанус, возникающий при более частом раздражении, когда каждый следующий импульс попадает в фазу сокращения (рис. 11). Таким образом, (в некоторых границах) между частотой импульсов возбуждения и амплитудой сокращения волокон ДЕ существует определенное соотношение: при небольшой частоте (например, 5-8 имп. в 1с)
54
Рис. П. Одиночноесокращение, зубчатыйи сплошной тетанускамбаловидпой мышцычеловека(по: Зимкин Н.В. и др., 1984). Верхняя кривая — сокращение мышцы, нижняя — отметкараздражения мышцы,справа указана частотараздражения
возникают одиночные сокращения, при увеличении частоты (15-20 имп. в 1с) — зубчатый тетанус, при дальнейшем нарастании частоты (25-60 имп. в 1 с) — гладкий тетанус. Одиночное сокращение — более слабое и менее утомительное, чем тетаническое. Зато тетанус обеспечивает в несколько раз более мощное, хотя и кратковременное сокращение мышечного волокна.
Сокращение целой мышцы зависит от формы сокращения отдельных ДЕ и их координации во времени. При обеспечении длительной, но не очень интенсивной работы, отдельные ДЕ сокращаются попеременно (рис. 12), поддерживая общее напряжение мышцы на заданном уровне (например, при беге на длинные и сверхдлинные дистанции). При этом отдельные ДЕ могут развивать как одиночные, так и тетанические сокращения, что зависит от частоты нервных импульсов. Утомление в этом случае развивается медленно, так как, работая по очереди, ДЕ в промежутках между активацией успевают восстанавливаться. Однако для мощного кратковременного усилия (например, поднятия штанги) требуется синхронизация активности отдельных ДЕ, т. е. одновременное возбуждение практически всех ДЕ. Это, в свою очередь, требует одновременной активации
55
Рис. 12. Различные режимы работы двигательных единиц (ДЕ)
соответствующих нервных центров и достигается в результате длительной тренировки. При этом осуществляется мощное и весьма утомительное тетаническое сокращение.
Амплитуда сокращения одиночного волокна не зависит от силы надпорогового раздражения (закон «Все или ничего»). В отличие от этого, при нарастании силы надпорогового раздражения сокращение целой мышцы постепенно растет до максимальной амплитуды.
Работа мышцы с небольшой нагрузкой сопровождается редкой частотой нервных импульсов и вовлечением небольшого числа ДЕ. В этих условиях, накладывая отводящие электроды на кожу над мышцей и используя усилительную аппаратуру, можно на экране осциллографа или с применением чернильной записи на бумаге зарегистрировать одиночные потенциалы действия отдельных Д Е. В случае же значительных напряжений потенциалы действия многих ДЕ алгебраически суммируются и возникает сложная интегрированная кривая записи электрической активности целой мышцы — электромиограмма (ЭМГ).
Форма ЭМГ отражает характер работы мышцы: при статических усилиях она имеет непрерывный вид, а при динамической работе — вид отдельных пачек импульсов, приуроченных, в основном, к начальному моменту сокращения мышцы и разделенных периодами «электрического молчания». Особенно хорошо ритмичность появления подобных пачек наблюдается у спортсменов при циклической работе (рис. 13). У маленьких детей и неадаптированных к такой работе лиц четких периодов отдыха не наблюдается, что отражает недостаточное расслабление мышечных волокон работающей мышцы.
Чем больше внешняя нагрузка и cилa сокращения мышцы, тем выше амплитуда ее ЭМГ. Это связано с увеличением частоты нервных импульсов, вовлечением большего числа ДЕ в мышце и синхронизацией
56
Рис. 13. Электромиограмма мышц-антагонистов при циклической работе
их активности. Современная многоканальная аппаратура позволяет производить одновременную регистрацию ЭМГ многих мышц на разных каналах. При выполнении спортсменом сложных движений можно видеть на полученных ЭМГ кривых не только характер активности отдельных мышц, но и оценить моменты и порядок их включения или выключения в различные фазы двигательных актов. Записи ЭМГ, полученные в естественных условиях двигательной деятельности, можно передавать к регистрирующей аппаратуре по телефону или радиотелеметрически. Анализ частоты, амплитуды и формы ЭМГ (например, с помощью специальных компьютерных программ) позволяет получить важную информацию об особенностях техники выполняемого спортивного упражнения и степени ее освоения обследуемым спортсменом.
По мере развития утомления при той же величине мышечного усилия амплитуда ЭМГ нарастает. Это связано с тем, что снижение сократительной способности утомленных ДЕ компенсируется нервными центрами вовлечением в работу дополнительных ДЕ, т. е. путем увеличения количества активных мышечных волокон. Кроме того, усиливается синхронизация активности ДЕ, что также повышает амплитуду суммарной ЭМГ.
-
МОРФОФУНКЦИОНАЛЬНЫЕ ОСНОВЫ МЫШЕЧНОЙ СИЛЫ
Движение является результатом взаимодействия внутренних и внешних сил, развиваемых в опорно-двигательном аппарате, — активных (возникающих при сокращении или напряжении мышцы во время ее возбуждения) и пассивных (упругое напряжение при растяжении мышцы, сопротивление мышцы и ее сухожилия).
Сила мышцы зависит отряда морфологических и физиологических факторов: количества и свойств мышечных волокон в мышце,
57
исходной длины мышцы, характера нервных импульсов, механических условий действия мышцы на кости скелета.
Сила мышцы является суммой силы отдельных ее мышечных волокон. Подсчитано, что 1 одиночное мышечное волокно икроножной мышцы развивает напряжение 100-200 мг, ІДЕ икроножной мышцы человека содержит около 2000 мышечных волокон и развивает напряжение 200-400 г, 1 икроножная мышца содержит около 1000 ДЕ, т. е. развивает напряжение 200-400 кг.
Большое значение имеет анатомическое строение мышцы. В параллельно-волокнистых и веретенообразных мышцах (камбаловидная мышца и др.) сила мышц тем больше, чем больше ее анатомический поперечник, т. е. площадь поперечного сечения целой мышцы. В перистых мышцах (двуглавая мышца и др.) физиологический поперечник, т. е. площадь поперечного сечения всех мышечных волокон гораздо больше, чем ее анатомический поперечник. В такой мышце упаковано значительно больше мышечных волокон и, соответственно, больше ее сила.
На силу сокращения мышцы влияет ее исходная длина, так как от нее. зависит возможное количество поперечных мостиков между актином и миозином. Предполагают, что в каждом цикле присоединения-отсоединения поперечных мостиков расходуется энергия 1 молекулы АТФ на 1 поперечный мостик. Следовательно, чем больше образуется в мышечном волокне актино-миозиновых мостиков, тем выше скорость расщепления АТФ, больше тяга сократительных белков и, соответственно, больше развиваемая мышцей сила.
Наибольшее количество актино-миозиновых контактов образуется при небольшом растяжении мышцы до некоторой оптимальной длины. При значительном растяжении саркомера нити актина далеко расходятся в стороны и практически не контактируют с расположенным в средней части саркомера миозином. В случае же резкого уменьшения длины саркомера нити актина в центре перекрывают друг друга, препятствуя контактам с миозином и также уменьшая число образуемых мостиков. В связи с этими особенностями взаимодействия сократительныхбелков наибольшая сила мышцы проявляется при некотором ее предварительном растяжении.
Одной из важнейших характеристик скелетных мышц, влияющих на силу сокращения, является состав (композиция) мышечных волокон. Различают 3 типа мышечных волокон — медленные неутомляемые (I типа), быстрые неутомляемые или промежуточные ( 11-а типа) и быстрые утомляемые (11-б типа).
Медленные волокна (1 типа), их обозначают также SO — Slow Oxydative (англ. — медленные окислительные) — это выносливые (неутомляемые) и легко возбудимые волокна, с богатым кровоснабжением, большим количеством митохондрий, запасов миоглобина и
58
с использованием окислительных процессов энергообразования (аэробные). Их, в среднем, у человека 50%. Они легко включаются в работу при малейших напряжениях мышц, очень выносливы, но не обладают достаточной силой. Чаще всего они используются при поддержании ненагрузочной статической работы, например, при сохранении позы.
Быстрые утомляемые волокна (11-б типа) или FG — Fast Glicolitic (быстрые гликолитические) используют анаэробные процессы энергообразования (гликолиз). Они менее возбудимы, включаются при больших нагрузках и обеспечивают быстрые и мощные сокращения мышц. Зато эти волокна быстро утомляются. Их примерно 30%. Волокна промежуточного типа (П-а) — быстрые неутомляемые, окислительные, их около 20%. В среднем, для разных мышц характерно различное соотношение медленных неутомляемых и быстрых утомляемых волокон. Так, в трехглавой мышце плеча преобладают быстрые волокна (67%) над медленными (33%), что обеспечивает скоростно-силовые возможности этой мышцы (рис. 14), а для более медленной и выносливой камбаловидной мышцы характерно наличие 84% медленных и всего 16% быстрых волокон (Салтан Б., 1979).
Однако, состав мышечных волокон в одной и той же мышце имеет огромные индивидуальные различия, зависящие от врожденных типологических особенностей человека. К моменту рождения человека его мышцы содержат лишь медленные волокна, но под влиянием нервной регуляции устанавливается в ходе онтогенеза генетически заданное индивидуальное соотношение мышечных волокон разного типа. По мере перехода от зрелого возраста к пожилому число быстрых волокон у человека заметно снижается и, соответственно, уменьшается мышечная сила. Например, наибольшее количество быстрых волокон в наружной головке 4-х главой мышцы бедра мужчины (около 59-63%) отмечается в возрасте 20-40 лет, а в возрасте 60-65 лет их число почти на 1/3 меньше (45%).
Рис. 14. Состав мышечных волокон в разных мышцах
Медленные — черным цветом; быстрые — серым
59
Количество тех или других мышечных волокон не изменяется в процессе тренировки. Возможно только нарастание толщины (гипертрофия) отдельных волокон, а также некоторое изменение свойств промежуточных волокон. При направленности тренировочного процесса на развитие силы происходит нарастание объема быстрых волокон, что и обеспечивает повышение силы тренируемых мышц.