Файл: Учебник для высших учебных заведений физической культуры Издание 2е, исправленное и дополненное.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.01.2024
Просмотров: 2170
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
12
железой внутренней секреции гипофизом и нервными клетками спинного и головного мозга. В настоящее время известно около сотни подобных веществ, которые являются осколками белков и, не вызывая сами возбуждения клеток, могут заметно изменять их функциональное состояние. Они влияют на сон, процессы обучения и памяти, на мышечный тонус (в частности, на позную асимметрию), вызывают обездвижение или обширные судороги мышц, обладают обезболивающим и наркотическим эффектом. Оказалось, что концентрация нейропептидов в плазме крови у спортсменов может превышать средний уровень у нетренированных лиц в 6-8 раз, повышая эффективность соревновательной деятельности. В условиях чрезмерных тренировочных занятий происходит истощение нейропептидов и срыв адаптации спортсмена к физическим нагрузкам.
2.3. РЕФЛЕКТОРНЫЙ МЕХАНИЗМ ДЕЯТЕЛЬНОСТИ НЕРВНОЙ СИСТЕМЫ
В деятельности нервной системы основным является рефлекторный механизм. Рефлекс — это ответная реакция организма на внешнее раздражение, осуществляемая с участием нервной системы.
Нервный путь рефлекса называется рефлекторной дугой. В состав рефлекторной дуги входят: 1) воспринимающее образование — рецептор, 2) чувствительный или афферентный нейрон, связывающий рецепторе нервными центрами, 3) промежуточные (или вставочные) нейроны нервных центров, 4) эфферентный нейрон, связывающий нервные центры с периферией, 5) рабочий орган, отвечающий на раздражение — мышца или железа.
Наиболее простые рефлекторные дуги включают всего две нервные клетки, однако множество рефлекторных дуг в организме состоят из значительного количества разнообразных нейронов, расположенных в различных отделах центральной нервной системы. Выполняя ответные реакции, нервные центры посылают команды к рабочему органу (например, скелетной мышце) через эфферентные пути, которые выполняют роль так называемых к а н а л о в прямой связи. В свою очередь, в ходе осуществления рефлекторного ответа или после него рецепторы, находящиеся в рабочем органе, и другие рецепторы тела посылают в центральную нервную систему информацию о результате действия. Афферентные пути этих сообщений — каналы обратной связи. Полученная информация используется нервными центрами для управления дальнейшими действиями, т. е. прекращением рефлекторной реакции, ее продолжением или изменением. Следовательно, основу
13
целостной рефлекторной деятельности составляет не отдельная рефлекторная дуга, а замкнутое рефлекторное кольцо, образованное прямыми и обратными связями нервных центров с периферией.
2.4. ГОМЕОСТАЗ
Внутренняя среда организма, в которой живут все его клетки, — это кровь, лимфа, межтканевая жидкость. Ее характеризует относительное постоянство — гомеостаз различных показателей, так как любые ее изменения приводят к нарушению функций клеток и тканей организма, особенно высокоспециализированных клеток центральной нервной системы. К таким постоянным показателям гомеостаза относятся температура внутренних отделов тела, сохраняемая в пределах 36-37° С, кислотно-основное равновесие крови, характеризуемое величиной рН = 7.4-7.35, осмотическое давление крови (7.6-7.8 атм.), концентрация гемоглобина в крови — 130-160 г. ּлֿ¹ и др.
Гомеостаз представляет собой не статическое явление, а динамическое равновесие. Способность сохранять гомеостаз в условиях постоянного обмена веществ и значительных колебаний факторов внешней среды обеспечивается комплексом регуляторных функций организма. Эти регуляторные процессы поддержания динамического равновесия получили название гомеокинеза.
Степень сдвига показателей гомеостаза при существенных колебаниях условий внешней среды или при тяжелой работе у большинства людей очень невелика. Например, длительное изменение рН крови всего на 0.1 -0.2 может привести к смертельному исходу. Однако, в общей популяции имеются отдельные индивиды, обладающие способностью переносить гораздо большие сдвиги показателей внутренней среды. У высококвалифицированных спортсменов-бегунов в результате большого поступления молочной кислоты из скелетных мышц в кровь во время бега на средние и длинные дистанции рН крови может снижаться до величин 7.0 и даже 6.9. Лишь несколько человек в мире оказались способными подняться на высоту порядка 8800 м над уровнем моря (на вершину Эвереста) без кислородного прибора, т. е. существовать и двигаться в условиях крайнего недостатка кислорода в воздухе и, соответственно, в тканях организма. Эта способность определяется врожденными особенностями человека — так называемой его генетической нормой реакции, которая даже для достаточно постоянных функциональных показателей организма имеет широкие индивидуальные различия.
14
2.5. ВОЗНИКНОВЕНИЕ ВОЗБУЖДЕНИЯ И ЕГО ПРОВЕДЕНИЕ 2.5.1. МЕМБРАННЫЕ ПОТЕНЦИАЛЫ
Мембрана клетки состоит из двойного слоя молекул липидов, повернутых «головками» наружу, а «хвостами» друг к другу. Между ними свободно плавают глыбы белковых молекул. Некоторые из них пронизывают мембрану насквозь. В части таких белков имеются, особые поры или ионные каналы, через которые могут проходить ионы, участвующие в образовании мембранных потенциалов (рис. I -А).
В возникновении и поддержании мембранного потенциала покоя основную роль играют два специальных белка. Один из них выполняет роль особогонатрий— калиевого насоса, который за счет энергии АТФ активно перекачивает натрий из клетки наружу, а калий внутрь клетки. В результате концентрация ионов калия становится внутри клетки выше, чем в омывающей клетку жидкости, а ионов натрия — выше снаружи.
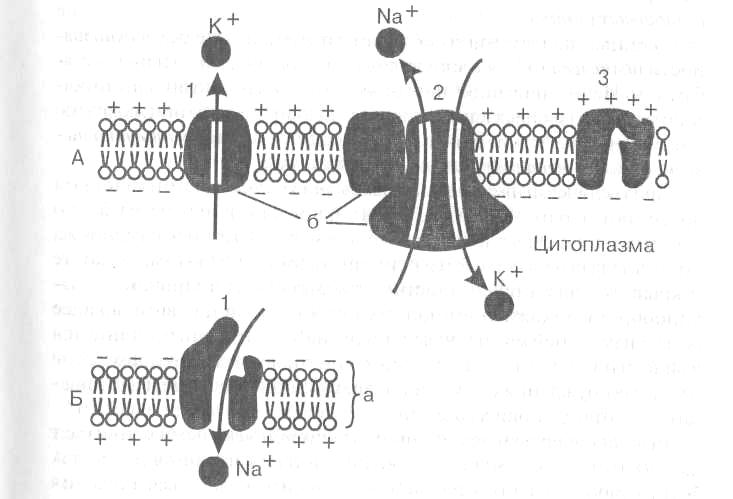
Рис. 1. Мембрана возбудимых клеток в покое (А) и при возбуждении (Б).
(По: Б.Альберте и др., 1986)
а — двойной слой липидов, б — белки мембраны.
На А: каналы «утечки калия» (1), «натрий-калиевый насос» (2)
и закрытый в покое натриевый канал (3).
На Б: открытый при возбуждении натриевый канал (1), вхождение ионов натрия в клетку и смена зарядов на наружной и внутренней стороне
мембраны.
15
Второй белок служит каналом утечки калия, через который ионы калия в силу диффузии стремятся выйти из клетки, где они содержатся в избытке. Ионы калия, выходя из клетки, создают положительный заряд на наружной поверхности мембраны. В результате внутренняя поверхность мембраны оказывается заряженной отрицательно по отношению к наружной. Таким образом, мембрана в состоянии покоя поляризована, т. е. имеется определенная разность потенциалов по обе стороны мембраны, называемая потенциалом покоя. Она равна для нейрона примерно минус 70 м В, для мышечного волокна — минус 90 мВ. Измеряют мембранный потенциал покоя, вводя тонкий кончик микроэлектрода внутрь клетки, а второй электрод помещая в окружающую жидкость. В момент прокола мембраны и вхождения микроэлектрода внутрь клетки на экране осциллографа наблюдают смещение луча, пропорциональное величине потенциала покоя.
В основе возбуждения нервных и мышечных клеток лежит повышение проницаемости мембраны для ионов натрия — открывание натриевых каналов. Внешнее раздражение вызывает перемещение заряженных частиц внутри мембраны и уменьшение исходной разности потенциалов по обе стороны или деполяризацию мембраны. Небольшие величины деполяризации приводят к открыванию части натриевых каналов и незначительному проникновению натрия внутрь клетки. Эти реакции являются подпороговыми и вызывают л и ш ь местные (локальные) изменения.
При увеличении раздражения изменения мембранного потенциала достигают порога возбудимости или критического уровня деполяризации — около 20 мВ, при этом величина потенциала покоя снижается примерно до минус 50 мВ. В результате открывается значительная часть натриевых каналов. Происходит лавинообразное вхождение ионов натрия внутрь клетки, вызывающее резкое изменение мембранного потенциала, которое регистрируется в виде потенциала действия. Внутренняя сторона мембраны в месте возбуждения оказывается заряженной положительно, а внешняя — отрицательно (рис. 1 -Б).
Весь этот процесс чрезвычайно кратковременный. Он занимает всего около
1-2 мс, после чего ворота натриевых каналов закрываются. К этому моменту достигает большой величины медленно нараставшая при возбуждении проницаемость для ионов калия. Выходящие из клетки ионы калия вызывают быстрое снижение потенциала действия. Однако окончательное восстановление исходного заряда продолжается еще некоторое время. В связи с этим в потенциале действия различают кратковременную высоковольтную часть — пик (или с п а й к) и длительные малые колебания — следовые потенциалы. Потенциалы действия мотонейронов имеют амплитуду пика около
16
100 мВ и длительность около 1.5 мс, в скелетных мышцах — амплитуда потенциала действия 120-130 мВ, адлительность2-3 мс.
В процессе восстановления после потенциалайействия работа натрий-калиевого насоса обеспечивает «откачку» излишних ионов натрия наружу и «накачивание» потерянных ионов калия внутрь, т. е. возвращение к исходной асимметрии их концентрации по обе стороны мембраны. На работу этого механизма тратится около 70% всей необходимой клетке энергии.
Возникновение возбуждения (потенциаладействия) возможно лишь при сохранении достаточного количества ионов натрия в окружающей клетку среде. Большие потери натрия организмом (например, с потом при длительной мышечной работе в условиях высокой температуры воздуха) могут нарушить нормальную деятельность нервных и мышечных клеток, снизив работоспособность человека. В условиях кислородного голодания тканей (например, при наличии большого кислородного долга во время мышечной работы) процесс возбуждения также нарушается из-за поражения (инактивации) механизма вхождения в клетку ионов натрия, и клетка становится невозбудимой. На процесс инактивации натриевого механизма влияет концентрация ионов Са
2.5.2. ПРОВЕДЕНИЕ ВОЗБУЖДЕНИЯ
Потенциалы действия (импульсы возбуждения) обладают способностью распространяться вдоль по нервным и мышечным волокнам.
В нервном волокне потенциал действия является очень сильным раздражителем для соседних участков волокна. Амплитуда потенциала действия обычно в 5-6 раз превышает пороговую величину деполяризации. Это обеспечивает высокую скорость и надежность проведения.
Между зоной возбуждения (имеющей на поверхности волокна отрицательный заряд и на внутренней стороне мембраны — положительный) и соседним не возбужденным участком мембраны нервного волокна (с обратным соотношением зарядов) возникают электрические токи — так называемые местные токи. В результате развивается деполяризация соседнего участка, увеличение его ионной проницаемости и появление потенциала действия. В исходной же зоне возбуждения восстанавливается потенциал покоя. Затем возбуждением охватывается следующий участок мембраны и т. д. Таким образом с помощью местных токов происходит распространение возбуждения на соседние участки нервного волокна, т.е. проведение нервного импульса. По мере проведения амплитуда потенциала действия не уменьшается, т. е. возбуждение не затухает даже при большой длине нерва.
17
В процессе эволюции с переходом от безмякотных нервных волокон к мякотным произошло существенное повышение скорости проведения нервного импульса. Для безмякотных волокон характерно непрерывное проведение возбуждения, которое охватывает последовательно каждый соседний участокнерва. Мякотные же нервы почти полностью покрыты изолирующей миелиновой оболочкой. Ионные токи в них могут проходить только в оголенных участках мембраны — перехватах Ранвье, лишенныхэтой оболочки. При проведении нервного им пульса возбуждение перескакивает от одного перехвата к другому и можетохватыватьдаже несколько перехватов. Такое проведение получило название сальтаторного (лат. saltus—прыжок). При этом повышается не только скорость, но и экономичность проведения. Возбуждение захватывает не всю поверхность мембраны волокна, а лишь небольшую ее часть. Следовательно, меньше энергии тратится на активный транспорт ионов через мембрану при возбуждении и в процессе восстановления.