Файл: Учебник для высших учебных заведений физической культуры Издание 2е, исправленное и дополненное.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.01.2024
Просмотров: 2173
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Проведение волны возбуждения от одного нейрона к другому через синапс происходит в большинстве нервных клеток химическим путем — с помощью медиатора, а медиатор содержится лишь в пресинаптической части синапса и отсутствует в постсинаптической мембране. Поэтому важной особенностью проведения возбуждения через синоптические контакты является одностороннее проведение нервных влияний, которое возможно лишь от пресинаптической мембраны к постсинаптической и невозможно в обратном направлении. В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление от афферентных нейронов к вставочным и затем к эфферентным — мотонейронам или вегетативным нейронам.
Большое значение в деятельности нервной системы имеет другая особенность проведения возбуждения через синапсы — замедленное проведение. Затрата времени на процессы, происходящие от момента подхода нервного импульса к пресинаптической мембране до появления в постсинаптической мембране потенциалов, называется синаптической задержкой. В большинстве центральных нейронов она составляет около 0.3 мс. После этого требуется еще время на развитие возбуждающего постсинаптического потенциала (ВПСП) и потенциала действия. Весь процесс передачи нервного импульса (от потенциала действия одной клетки до потенциала действия следующей клетки) через один синапс занимает примерно 1.5 мс. При утомлении, охлаждении и ряде других воздействий длительность синаптической задержки возрастает. Если же для осуществления какой-либо реакции требуется участие большого числа нейронов (многих сотен и даже тысяч), то суммарная величина задержки проведения по нервным центрам может составить десятые доли секунды и даже целые секунды.
При рефлекторной деятельности общее время от момента нанесения внешнего раздражения до появления ответной реакции организма—так называемое скрытое или латентное время рефлекса определяется в основном длительностью проведения через синапсы. Величина латентного времени рефлекса служит важным показателем функционального состояния нервных центров. Измерение латентного времени простой двигательной реакции человека на внешний сигнал широко используется в практике для оценки функционального состояния ЦНС (рис. 3).
23
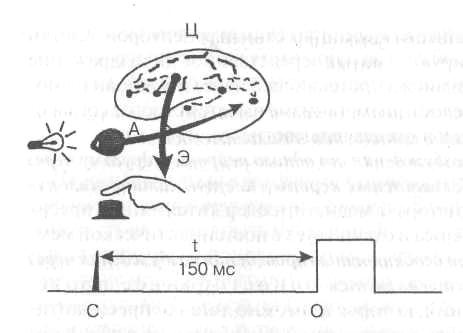
Рис. 3. Схема измерения
времени двигательной
реакции
А — афферентные,
Э — эфферентные и
Ц — центральные пути;
С— отметка светового
сигнала, О — отметка нажима
кнопки,
tISOmc — время реакции.
3.3.2. СУММАЦИЯ ВОЗБУЖДЕНИЯ
В ответ на одиночную афферентную волну, идущую от рецепторов к нейронам, в пресинаптической части синапса освобождается небольшое количество медиатора. При этом в постсинаптической мембране нейрона обычно возникает ВПСП — небольшая местная деполяризация. Для того, чтобы общая по всей мембране нейрона величина ВПСП достигала порога возникновения потенциала действия, требуется суммация на мембране клетки многих подпороговых ВПСП. Лишь в результате такой суммации возбуждения возникает ответ нейрона. Различают пространственную и временную суммацию.
Пространственная суммация наблюдается в случае одновременного поступления нескольких импульсов в один и тот же нейрон по разным пресинаптическим волокнам. Одномоментное возбуждение синапсов в различных участках мембраны нейрона повышает амплитуду суммарного ВПСП до пороговой величины. В результате возникает ответный импульс нейрона и осуществляется рефлекторная реакция. Например, для получения ответа двигательной клетки спинного мозга обычно требуется одновременная активация 50-100 афферентных волокон от соответствующих периферических рецепторов..
Временная суммация происходит при активации одного и того же афферентного пути серией последовательных раздражений. Если интервалы между поступающими импульсами достаточно коротки и ВПСП нейрона от предыдущих раздражений не успевают затухать, то последующие ВПСП накладываются друг на друга, пока деполяризация мембраны нейрона не достигнет критического уровня для возникновения потенциала действия. Таким способом даже слабые раздражения через некоторое время могут вызывать ответные реакции организма (например, чихание и кашель в ответ на слабые раздражения слизистой оболочки дыхательных путей).
24
3.3.3. ТРАНСФОРМАЦИЯ И УСВОЕНИЕ РИТМА
Характер ответного разряда нейрона зависит не только от свойств раздражителя, но и от функционального состояния самого нейрона (его мембранного заряда, возбудимости, лабильности). Нервные клетки обладают свойством изменять частоту передающихся импульсов
, т.е. свойством трансформации ритма.
При высокой возбудимости нейрона (например, после приема кофеина) может возникать учащение импульсации (мультипликация ритма), а при низкой возбудимости (например, при утомлении) происходит урежение ритма, так как несколько приходящих импульсов должны суммироваться, чтобы наконец достичь порога возникновения потенциала действия. Эти изменения частоты импульсации могут усиливать или ослаблять ответные реакции организма на внешние раздражения.
При ритмических раздражениях активность нейрона может настроиться на ритм приходящих импульсов, т. е. наблюдается явление усвоения ритма (Ухтомский А. А., 1928). Развитие усвоения ритма обеспечивает сонастройку активности многих нервных центров при управлении сложными двигательными актами, особенно это важно для поддержания темпа циклических упражнений.
3.3.4. СЛЕДОВЫЕ ПРОЦЕССЫ
После окончания действия раздражителя активное состояние нервной клетки или нервного центра обычно продолжается еще некоторое время. Длительность следовых процессов различна: небольшая в спинном мозге (несколько секунд или минут), значительно больше в центрах головного мозга (десятки минут, часы или даже дни) и очень большая в коре больших полушарий (до нескольких десятков лет).
Поддерживать явное и кратковременное состояние возбуждения в нервном центре могут импульсы, циркулирующие по замкнутым цепям нейронов. Значительно сложнее по природе длительно сохраняющиеся скрытые следы. Предполагают, что длительное сохранение в нервной клетке следов со всеми характерными свойствами раздражителя основано на изменении структуры составляющих клетку белков и на перестройке синаптических контактов.
Непродолжительные импульсные последействия (длительностью до 1 часа) лежат в основе так называемой кратковременной памяти, а длительные следы, связанные со структурными и биохимическими перестройками в клетках, — в основе формирования долговременной памяти.
25
3.4. КООРДИНАЦИЯ ДЕЯТЕЛЬНОСТИ ЦНС
Процессы координации деятельности ЦНС основаны на согласовании двух нервных процессов — возбуждения и торможения. Торможение является активным нервным процессом, который предупреждает или угнетает возбуждение.
3.4.1. ЗНАЧЕНИЕ ПРОЦЕССА ТОРМОЖЕНИЯ В ЦНС
Явление торможения в нервных центрах было впервые открыто И. М. Сеченовым в 1862 г. Значение этого процесса было рассмотрено им в книге «Рефлексы головного мозга» (1863).
Опуская лапку лягушки в кислоту и одновременно раздражая некоторые участки головного мозга (например, накладывая кристаллик поваренной соли на область промежуточного мозга), И. М. Сеченов наблюдал резкую задержку и даже полное отсутствие «кислотного» рефлекса спинного мозга (отдергивания лапки). Отсюда он сделал заключение, что одни нервные центры могут существенно изменять рефлекторную деятельность в других центрах, в частности вышележащие нервные центры могут тормозить деятельность нижележащих. Описанный опыт вошел в историю физиологии под названием Сеченовское торможение.
Тормозные процессы —необходимый компонент в координации нервной деятельности. Во-первых, процесс торможения ограничивает распространение возбуждения на соседние нервные центры, чем способствуетего концентрации в необходимых участках нервной системы. Во-вторых, возникая в одних нервных центрах параллельно с возбуждением других нервных центров, процесс торможения тем самым выключает деятельность ненужных в данный момент органов. В-третьих, развитие торможения в нервных центрах предохраняет их от чрезмерного перенапряжения при работе, т. е. играет охранительную роль.
3.4.2. ПОСТСИНАПТИЧЕСКОЕ И ПРЕСИНАПТИЧЕСКОЕ ТОРМОЖЕНИЕ
Процесс торможения, в отличие от возбуждения, не может распространяться по нервному волокну — это всегда местный процесс в области синаптических контактов. По месту возникновения различают пресинаптическое и постсинаптическое торможение.
Постсинаптическое торможение — это тормозные эффекты, возникающие в постсинаптической мембране. Чаще всего этот вид торможения связан с наличием в ЦНС специальных тормозных нейронов. Они представляют собой особый тип вставочных нейронов, у которых окончания аксонов выделяют тормозный
26
медиатор. Одним из таких медиаторов является гамма-аминомасляная кислота (ГАМК).
Нервные импульсы, подходя к тормозным нейронам, вызывают в них такой же процесс возбуждения, как и в других нервных клетках. В
ответ по аксону тормозной клетки распространяется обычный потенциал действия. Однако, в отличие от других нейронов, окончания аксона при этом выделяют не возбуждающий, а тормозной медиатор. В результате тормозные клетки тормозят те нейроны, на которых оканчиваются их аксоны.
К специальным тормозным нейронам относятся клетки Рэншоу в спинном мозге, клетки Пуркинье мозжечка, корзинчатые клетки в промежуточном мозге и др. Большое значение, например, тормозные клетки имеют при регуляции деятельности мышц-антагонистов: приводя к расслаблению мышц антагонистов, они облегчают тем самым одновременное сокращение мышц-агонистов (рис.4).
Клетки Рэншоу участвуют в регуляции уровня активности отдельных мотонейронов спинного мозга. При возбуждении мотонейрона импульсы поступают по его аксону к мышечным волокнам и одновременно по коллатералям аксона — к тормозной клетке Рэншоу. Аксоны последней «возвращаются» к этому же нейрону, вызывая его торможение. Чем больше возбуждающих импульсов посылает мотонейрон на периферию (а значит, и к тормозной клетке), тем сильнее это возвратное торможение (разновидность постсинаптического торможения). Такая замкнутая система действует
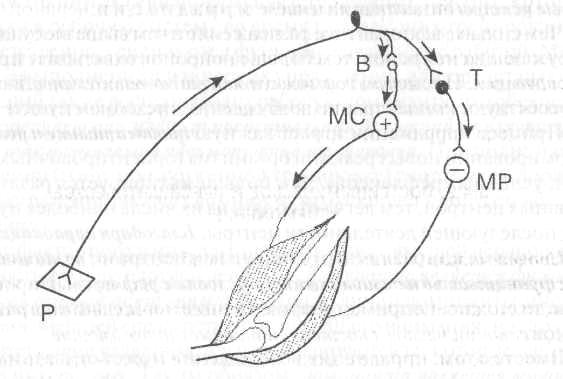
Рис. 4. Участие тормозной клет
ки в регуляции мышц-антагонистов
В и Т— возбуждающий и тормозный нейроны. Возбуждение ('+) мотонейрона мышцы-сгибателя (МС) и торможение (—) мотонейрона мышцы-разгибателя (МР). Р — кожный рецептор.