ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 22.04.2024
Просмотров: 377
Скачиваний: 1
СОДЕРЖАНИЕ
7.4 Энергообеспечение динамической работы при подтягивании. 27
7.5 Оценка уровня развития силовых способностей по внешним признакам. 31
7.1 Мышцы, производящие подъём/опускание туловища.
7.2 Строение мышечных волокон и механизм мышечных сокращений
7.2.1 Строение и химический состав скелетных мышц
7.2.2 Механизм мышечного сокращения.
7.2.3 Изменение величины силы в фазе подъёма
7.3 Изменения в мышечных волокнах под влиянием различных тренировочных воздействий.
7.3.1 Особенности различных типов мышечных волокон
7.3.2 Увеличение количества миофибрилл в быстрых мышечных волокнах
1 Подтягивание с большими грузами.
3 Интервальная тренировка с отягощением.
7.3.3 Увеличение количества митохондрий в быстрых мышечных волокнах
1 Подтягивание со спрыгиванием.
2 Подтягивание в сверхнизком темпе.
7.3.4 Параллельное увеличение количества митохондрий и миофибрилл в быстрых мышечных волокнах
7.3.5 Увеличение количества миофибрилл в медленных мышечных волокнах
1 Увеличение силы мышц-сгибателей пальцев.
2 Развитие силы ммв мышц, выполняющих подъём туловища.
7.3.6 Увеличение количества митохондрий в медленных мышечных волокнах
7.3.7 Схема изменений в мышечных волокнах под воздействием нагрузки.
7.4 Энергообеспечение динамической работы при подтягивании.
7.4.1 Энергообеспечение динамической работы при подтягивании в оптимальном соревновательном темпе
7.4.2 Энергообеспечение динамической работы при подтягивании в низком темпе
7.4.3 Энергообеспечение динамической работы при подтягивании в повышенном темпе
7.4.4 Энергообеспечение динамической работы при подтягивании в максимальном темпе
7.5 Оценка уровня развития силовых способностей по внешним признакам.
7.6 Динамические силовые способности и результат в подтягивании.
7.7 Условия для повышения динамических силовых способностей
7.10.1 Особенности построения тренировочной нагрузки.
7.10.5 Краткое описание тренировочного процесса с применением повторно-серийного метода.
7.2.3 Изменение величины силы в фазе подъёма
Количество поперечных мостиков, связывающих актиновые и миозиновые нити, а, следовательно, и развиваемая мышечная сила согласно теории скользящих нитей, зависит от степени перекрытия толстых и тонких нитей, а значит, и от длины саркомера или мышцы. Максимум развиваемой изолированной мышцей силы будет наблюдаться при длине, примерно соответствующей состоянию покоя. При уменьшении длины сила уменьшается из-за того, что актиновые и миозиновые нити начинают мешать друг другу, а при растяжении мышцы до большей, чем в покое, длины сила уменьшается из-за того, что нити актина оказываются вытянутыми из миозиновых пучков. При этом только часть головок миозина может присоединиться к актину [31].
В реальных условиях мышцы, прикрепляющиеся к костям, вызывают движение рабочих звеньев тела. С движением рабочего звена меняется угол в суставе и, следовательно, длина обслуживающих данное сочленение мышц и угол подхода их к месту прикрепления на кости. При этом увеличиваются или уменьшаются плечо и момент силы мышц, что, в свою очередь, изменяет механические условия их работы. Эти условия могут быть выгодными, когда силовой потенциал используется полностью, и невыгодными, когда максимальное напряжение мышц используется только частично [18].
Сила, проявляемая в наименее целесообразной с механической точки зрения фазе движения, часто составляет не более 50-60% (рисунок 7.7) от силы в наиболее целесообразной его фазе [23].
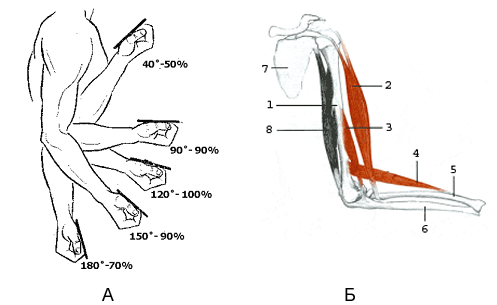
Рисунок 7.7А - динамика максимальной силы при сгибании руки в локтевом суставе (по Платонову В.Н.).
Б - рисунок, иллюстрирующий сгибание руки в локтевом суставе (по [27]).
1 – плечевая кость; 2 - двуглавая мышца плеча; 3 - плечевая мышца; 4 - плечелучевая мышца; 5 – лучевая кость; 6 – локтевая кость; 8 - трёхглавая мышца плеча (разгибатель локтевого сустава)
При подтягивании на перекладине наименее благоприятным является верхний участок траектории движения в фазе подъёма, который недостаточно подготовленные спортсмены проходят с большим трудом, особенно в конечной части выполнения упражнения.
В начале подтягиваний, когда силовые способности спортсмена находятся на должном уровне, он может проходить проблемный участок по инерции – за счёт набранной ранее скорости. По мере накопления усталости силовые способности спортсмена снижаются настолько, что он уже не может набрать необходимую скорость. Поэтому в случае недостаточного уровня развития силы спортсмен «зависает» на верхнем участке траектории движения, затрачивая на его прохождение неоправданно большое количество энергии.
На рисунке 7.8 приведены графики зависимости резерва силовых способностей от высоты подъёма для 6 человек – 5 спортсменов (КМС и МС по полиатлону) и одного человека, не занимающегося спортом. Резервом силы в данном случае считается сила, которую способен развить испытуемый в какой-либо точке траектории движения сверх веса своего тела. Поскольку рост (а значит, и высота подъёма) и вес тела спортсменов различен, их силовые способности следует выражать в относительных единицах. Так, высота подъёма из исходного положения до уровня грифа принята за 100%, а резерв силы выражен в долях веса тела каждого из участников эксперимента.
В нижней части траектории движения (от 0 до 30 %) характер изменения резерва силы может быть различным, но, начиная примерно с 30% от высоты подъёма, резерв силы непрерывно снижается, что является экспериментальным подтверждением тезиса о проблемности верхнего участка траектории.
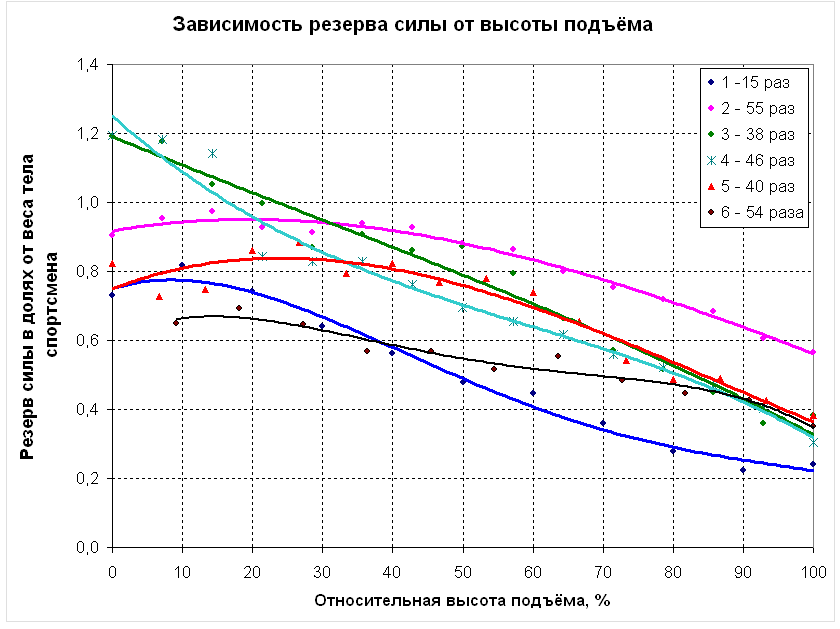
Рисунок 7.8Изменение относительного резерва силы в фазе подъёма.
Взаимосвязь относительного резерва силы и результата в подтягивании имеет сложный характер. С одной стороны, испытуемый, не занимающийся подтягиванием (кривая 1), имеет более низкие показатели резерва силы в верхней части траектории движения по сравнению с квалифицированными полиатлонистами. Но с другой стороны, спортсмены, имеющие различные результаты в подтягивании (от 38 до 54 раз – кривые 3-6), показали в эксперименте практически равные значения относительного резерва силы на верхнем участке траектории движения. В то же время у спортсменов, имеющих практически равные результаты в подтягивании (54 и 55 раз – кривые 6 и 2), наблюдаются существенно отличающиеся показатели резерва силы на уровне грифа – соответственно 35% и 56% от веса тела. Это можно объяснить тем, что при достижении определённого уровня квалификации в подтягивании (предположительно 35-40 раз) первостепенное значение имеет не столько величина резерва силы, которая зависит от анаэробных возможностей мышц спортсмена, сколько скорость его расходования при выполнении подтягиваний, которая в большей степени определяется аэробными возможностями мышц.
Так, для спортсмена 2, резерв силы которого на уровне грифа перекладины составляет 56% от веса тела, средняя скорость расходования этого резерва будет составлять 56/55=1,02 % от веса тела в расчёте на одно подтягивание. Для спортсмена 6, резерв силы которого в той же точке составляет 35% от веса тела, скорость его снижения будет равна 35/54=0,65% от веса тела в расчёте на одно подтягивание. Следовательно, спортсмен 6, обладающий существенно меньшим резервом силы, выполняет подтягивания более экономично.
В параграфе 2.4.2 мы рассматривали соотношение между силой и скоростью мышечного сокращения (кривая Хилла), отметив, что скорость сокращения мышцы нелинейно снижается с увеличением нагрузки. Ненагруженная мышца сокращается с максимальной скоростью, но эта скорость зависит от типа мышечных волокон. Чем быстрее (при участии АТФ) происходит циклическое взаимодействие миозиновых мостиков с актином, тем выше скорость скольжения. АТФазная активность миозина в быстрых мышечных волокнах выше, чем в медленных, поэтому поперечные мостики волокон типа IIА иIIВ будут быстрее расщеплять АТФ и взаимодействовать с актином.
В условиях противодействия нагрузке максимальная скорость сокращения мышцы будет тем ниже, чем выше нагрузка. В случае если нагрузка не превышает предельные возможности спортсмена, он может произвольно менять скорость мышечного сокращения (например, скорость в фазе подъёма туловища). Так, когда все мышечные волокна участвуют в поднимании груза, относительная нагрузка на каждое мышечное волокно меньше (а скорость их сокращения больше), чем в том случае, когда активна лишь часть волокон. Таким образом, можно увеличить скорость укорочения мышцы при одной и той же нагрузке за счёт вовлечения дополнительных двигательных единиц [31].
Теоретически увеличение максимальной произвольной силы спортсмена должно приводить к улучшению результата в подтягивании за счёт относительного снижения нагрузки на мышцы и возможности подключения дополнительных двигательных единиц в ходе выполнения подтягиваний. Но на практике это часто происходит только в том случае, если длительность подтягиваний не превышает 1-2 минуты. И связано это с тем, что увеличение максимальной силы требует проведения соответствующих тренировок с большими отягощениями, которые направлены на увеличение количества миофибрилл и повышение концентрации креатинфосфата в быстрых мышечных волокнах. Но тренировки, состоящие из подходов с большими отягощениями, отрицательно сказываются на длительности виса. Получается замкнутый круг – увеличение одной способности происходит за счёт снижения другой, разорвать который можно только за счёт разумного сочетания тренировок, направленных на увеличение количества миофибрилл и тренировок, направленных на повышение числа митохондрий в рабочих мышцах спортсмена.
7.3 Изменения в мышечных волокнах под влиянием различных тренировочных воздействий.
В соответствии с теорией функциональных систем П.К.Анохина поведение любой системы (в том числе и двигательной системы спортсмена) подчинено получению определённого полезного результата, а недостаточность этого результата может полностью реорганизовать систему и сформировать новую, с более совершенным взаимодействием компонентов, дающим достаточный результат. Таким образом, результат является неотъемлемым и решающим компонентом системы, инструментом, создающим упорядоченное взаимодействие между всеми другими её компонентами [36].
Но каким образом спортсмен (как личность) может объяснить своему организму (как мешку с костями) что он хочет подтянуться 50 раз за 4 минуты, в то время как тот может только 15 раз за полторы? Видимо, ему придётся разговаривать со своим организмом на понятном тому языке. Поскольку исполнительным органом двигательной системы являются сократительные структуры мышечных клеток, то и язык этот должен быть языком, понятным для мышц. Мышцы же (в естественных условиях) понимают только один язык – язык физической нагрузки, для выполнения которой они, собственно говоря, и нужны.
Определённым образом задавая нагрузку, мы можем вызвать желательные изменения физических возможностей спортсмена. Чтобы эти изменения позволили спортсмену со временем перейти от 15 к 50 подтягиваниям, нагрузка должна указывать мышечной системе (понятным ей способом) направление проведения необходимых изменений, т.е. нагрузка должна быть целенаправленной.
Экспериментальный – методом проб и ошибок - отбор упражнений наряду с теоретическим обоснованием их применения позволяет подобрать такие упражнения, которые обеспечивают необходимую направленность тренировочного процесса.
Но мало задать целенаправленную нагрузку, нужно ещё и добиться от организма, чтобы тот производил «модернизацию» двигательной системы до тех пор, пока поставленная спортсменом цель не будет достигнута. Поэтому нагрузка, во-первых, должна быть такой, чтобы организм напрягался при её выполнении, т.е. она должна быть развивающей. А во-вторых, для того чтобы обеспечить непрерывное движение к поставленной цели, нагрузка должна периодически изменяться, увеличиваясь и не давая организму «расслабляться» до тех пор, пока не будет получен необходимый результат.
Таким образом, применение цепочки связанных одной целью нагрузок приводит к тому, что, достигнув определённого (промежуточного) результата, организм сразу начинает «беспокоиться» по поводу следующего результата, что в конечном итоге и приводит к достижению главной цели. А значит, цепочка целенаправленных нагрузок будет стимулировать двигательную функциональную систему произвести такие перестройки в организме спортсмена, которые и позволят со временем перейти от 15 к 50 подтягиваниям.
Но поскольку нужный результат может быть получен различными способами (это обсуждалось в п. 5.2), коротко рассмотрим эти способы в привязке к изменениям аэробных и силовых возможностей мышц, т.е. используя «язык митохондрий и миофибрилл».
7.3.1 Особенности различных типов мышечных волокон
Итак, миофибриллы представляют собой сократительный аппарат мышечных клеток. Их количество определяет величину мышечной силы. Митохондрии отвечают за окислительные возможности мышц, т.е. за способность ресинтеза АТФ аэробным способом.
При подтягивании на перекладине требуется многократное проявление силы заданного уровня, причём этот уровень таков, что для подъёма туловища недостаточно включения только низкопороговых двигательных единиц. Необходимо участие высокопороговых ДЕ, которые в основном состоят из так называемых быстрых мышечных волокон (БМВ), осуществляющих ресинтез АТФ анаэробным способом. В связи с этим тренировка в подтягивании должна строиться как по пути увеличения силовых возможностей мышц (за счёт увеличения количества миофибрилл), так и по пути повышения окислительных возможностей мышц, участвующих в подъёме туловища и удержании хвата (за счёт увеличения количества митохондрий).
Напомним, что:
Медленные (окислительные) мышечные волокна (ММВ - тип I) характеризуются высокой активностью окислительных ферментов, наличием большого количества митохондрий, низкой активностью миозин-АТФ-азы – фермента, способствующего образованию поперечных мостиков при мышечном сокращении. Медленные волокна имеют богатую капиллярную сеть, а повышенное содержание миоглобина облегчает транспорт кислорода к митохондриям внутри мышечной клетки. Перечисленные особенности объясняют использование медленными мышечными волокнами аэробного пути энергообеспечения и их способность к выполнению длительной работы преимущественно аэробного характера
Быстрые окислительно-гликолитические мышечные волокна (тип IIA), обладающие мощной анаэробной системой энергопродукции, приспособлены также и к выполнению достаточно интенсивной аэробной работы. С функциональной точки зрения эти волокна рассматриваются как промежуточные между медленными (тип I) и быстрыми гликолитическими (тип IIВ).
Быстрые гликолитические мышечные волокна отличаются высокой активностью АТФ-азы и ферментов гликолиза и низкой активностью окислительных ферментов. Слабо развитая капиллярная сеть, малое количество митохондрий и миоглобина в их составе означает, что такие волокна не обладают большой выносливостью, но способны выполнять мощные и быстрые (хотя и относительно кратковременные) мышечные сокращения.