Файл: Контрольная работа по дисциплине генетика вариант 8 Выполнил(а) студент(ка) 2 курса, зоо728о ип.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 26.10.2023
Просмотров: 122
Скачиваний: 2
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Министерство сельского хозяйства Российской Федерации
ФГБОУ ВО «Вологодская государственная молочнохозяйственная академия имени Н. В. Верещагина»
Факультет ветеринарной медицины и биотехнологий
Направление подготовки «Ветеринария»
Форма обучения очно-заочная
Кафедра зоотехнии и биологии
Контрольная работа
по дисциплине
«ГЕНЕТИКА»
вариант 8
Выполнил(а)
студент(ка) 2 курса,
ЗОО-728-О ИП
шифр 2117048 Ратушиной К. В.
Проверила,
к. с.-х. н, доцент Н.Ю.Литвинова
Вологда-Молочное
2023 г
Содержание
Содержание 3
Вопрос №3. Вклад в развитии генетики отечественных исследователей 4
Вопрос № 10. Генетический код, его сущность и основные свойства (триплетность, неперекрываемость, универсальность) 9
Вопрос №58. Повторяемость хозяйственно-полезных признаков, значение и использование 13
Задача № 65 16
Задача № 76 16
Задача № 99 16
Список использованной литературы 18
Вопрос №3. Вклад в развитии генетики отечественных исследователей
Если не считать опытов по гибридизации растений в XVIII в., первые работы по генетике в России были начаты в начале XX в. как на опытных сельскохозяйственных станциях, так и в среде университетских биологов, преимущественно тех, кто занимался экспериментальной ботаникой и зоологией.
После революции и гражданской войны 1917—1922 гг. началось стремительное организационное развитие науки. К концу 1930-х годов в СССР была создана обширная сеть научно-исследовательских институтов и опытных станций (как в Академии наук СССР, так и во Всесоюзной академии сельскохозяйственных наук имени Ленина (ВАСХНИЛ)), а также вузовских кафедр генетики. Признанными лидерами направления были Н. И. Вавилов, Н. К. Кольцов, А. С. Серебровский, С. С. Четвериков и др. В СССР издавали переводы трудов иностранных генетиков, в том числе Т. Х. Моргана, Г. Мёллера, ряд генетиков участвовали в международных программах научного обмена. Американский генетик Г. Мёллер работал в СССР (1934—1937), советские генетики работали за границей. Н. В. Тимофеев-Ресовский — в Германии (с 1925 г.), Ф. Г. Добржанский — в США (с 1927 г.) [1, c. 3-4].
В 1930-е гг. в рядах генетиков и селекционеров наметился раскол, связанный с энергичной деятельностью Т. Д. Лысенко и И. И. Презента. По инициативе генетиков был проведён ряд дискуссий (наиболее крупные — в 1936 и 1939 г.), направленных на борьбу с подходом Лысенко.
На рубеже 1930—1940-х гг. в ходе так называемого Большого террора большинство сотрудников аппарата ЦК ВКП (б), курировавших генетику, и ряд видных генетиков были арестованы, многие расстреляны или погибли в тюрьмах (в том числе, Н. И. Вавилов). После войны дебаты возобновились с новой силой. Генетики, опираясь на авторитет международного научного сообщества, снова попытались склонить чашу весов в свою сторону, однако с началом холодной войны ситуация значительно изменилась. В 1948 году на августовской сессии ВАСХНИЛ Т. Д. Лысенко, пользуясь поддержкой И. В. Сталина, объявил генетику лженаукой. Лысенко воспользовался некомпетентностью партийного руководства наукой, «пообещав партии» быстрое создание новых высокопродуктивных сортов зерна («ветвистая пшеница») и др. С этого момента начался период гонений на генетику, который получил название лысенковщины и продолжался вплоть до снятия Н. С. Хрущева с поста генерального секретаря ЦК КПСС в 1964 г [1, c. 4-5].
Лично Т. Д. Лысенко и его сторонники получили контроль над институтами отделения биологии АН СССР, ВАСХНИЛ и вузовскими кафедрами. Были изданы новые учебники для школ и вузов, написанные с позиций «Мичуринской биологии». Генетики вынуждены были оставить научную деятельность или радикально изменить профиль работы. Некоторым удалось продолжить исследования по генетике в рамках программ по изучению радиационной и химической опасности за пределами организаций, подконтрольных Т. Д. Лысенко и его сторонникам.
После открытия и расшифровки структуры ДНК, физической базы генов (1953 г.), с середины 1960-х г. началось восстановление генетики. Министр просвещения РСФСР В. Н. Столетов инициировал широкую дискуссию между лысенковцами и генетиками, в результате было опубликовано много новых работ по генетике. В 1963 г. вышел в свет университетский учебник М. Е. Лобашева «Генетика», выдержавший впоследствии несколько изданий. Вскоре появился и новый школьный учебник Общая биология под редакцией Ю. И. Полянского, используемый, наряду с другими, и по сей день [4, c. 10].
Николай Иванович Вавилов (13 (25) ноября 1887, Москва, Российская империя — 26 января 1943, Саратов, РСФСР, СССР) — российский и советский учёный-генетик, ботаник, селекционер, географ, академик АН СССР, АН УССР и ВАСХНИЛ. Президент (1929—1935), вице-президент (1935—1940) ВАСХНИЛ, президент Всесоюзного географического общества (1931—1940), основатель (1920) и бессменный до момента ареста директор Всесоюзного института растениеводства (1930—1940), директор Института генетики АН СССР (1930—1940), член Экспедиционной комиссии АН СССР, член коллегии Наркомзема СССР, член президиума Всесоюзной ассоциации востоковедения. В 1926—1935 годах член Центрального исполнительного комитета СССР, в 1927—1929 — член Всероссийского Центрального Исполнительного Комитета.
Организатор и участник ботанико-агрономических экспедиций, охвативших большинство континентов (кроме Австралии и Антарктиды), в ходе которых выявил древние очаги формообразования культурных растений. Создал учение о мировых центрах происхождения культурных растений. Обосновал учение об иммунитете растений, открыл закон гомологических рядов в наследственной изменчивости организмов. Внёс существенный вклад в разработку учения о биологическом виде. Под руководством Вавилова была создана крупнейшая в мире коллекция семян культурных растений. Он заложил основы системы государственных испытаний сортов полевых культур. Сформулировал принципы деятельности главного научного центра страны по аграрным наукам, создал сеть научных учреждений в этой области [4, c. 11].
Николай Константинович Кольцов (3 (15) июля 1872, Москва — 2 декабря 1940 года, Ленинград) — выдающийся русский биолог, автор идеи матричного синтеза.
Начиная свою работу в период расцвета описательной биологии и первых шагов экспериментальной биологии, Кольцов тонко чувствовал тенденции развития биологии и рано осознал значение экспериментального метода. Он проповедовал необходимость экспериментального подхода во всех областях биологии и предсказал его использование даже в эволюционном учении (не противопоставляя экспериментальные методы описательным). Речь шла не о простом биологическом эксперименте, а об использовании методов физики и химии. Кольцов не раз подчеркивал огромное значение для биологии открытия новых форм лучистой энергии, в частности рентгеновских и космических лучей, писал о применении радиоактивных веществ. Чтобы изучить организм в целом, надо использовать все современные знания в области физической и коллоидной химии, необходимо изучать внутри клетки мономолекулярные слои и их роль в разнообразных превращениях веществ. "Биологи ждут, когда эти методы (рентгеноструктурного анализа) будут усовершенствованы настолько, что можно будет при их помощи изучить кристаллическую структуру внутриклеточных скелетных, твердых структур белкового и иного характера". Эта мысль явилась пророческой и реально осуществилась в открытии методом рентгеноструктурного анализа строения молекулы ДНК. Пророческой оказалась и другая идея Кольцова, в которой он также шел от биологии к химии. Исходя из развиваемого им представления, что каждая сложная биологическая молекула возникает из подобной ей уже существующей молекулы, он предсказал, что химики пойдут по пути создания новых молекул в растворах, содержащих необходимые составные части сложных молекул, путем внесения в них затравок готовых молекул той же структуры. Он писал: "Я думаю, что только таким способом удастся синтезировать in vitro белки, и при том не какие-нибудь, а определенные, т. е. синтез которых заранее намечается" [2, c. 20].
Одними из первых опыты в данной области проделали русские ученые Георгий Адамович Надсон и его молодой сотрудник Григорий Семенович Филиппов, работавшие в Институте рентгенологии и радиологии.
Они получили мутации у дрожжей и низших грибов под действием радия и рентгенных лучей. В результате экспериментов вырастали колонии, отличавшиеся величиной, формой, окраской, дрожжевые клетки изменяли свои биохимические свойства. Таким образом была открыта способность радиации вызывать наследственные изменения.
В середине 30-х годов была сформулирована теория, описывающая кинетические зависимости ни активирующего и мутагенного эффекта ионизирующих излучений - так называемая "теория мишени". Важнейшие эксперименты, ставшие основой этой теории, были проведены в период 1931 - 1937 гг. несколькими исследователями, среди которых был и Николай Владимирович Тимофеев-Ресовский, ставший одним из основателей количественной биофизики ионизирующих излучений [2, c. 21].
Тимофеев-Ресовский развивал идеи Н. Кольцова, предполагавшего, что молекулярные наследственные структуры образуются посредством матричного синтеза. Он вел исследования по биофизическому анализу мутационного процесса, впоследствии приведшие к формированию молекулярной биологии как новой синтетической дисциплины. Тимофеев-Ресовский показал, что мутационные изменения затрагивают относительно ограниченную группу атомов в хромосоме. Это открытие впервые переводило мутационный процесс на молекулярный уровень понимания.
Первым, кто обратил внимание на мутации, вызванные химическими веществами, был Н.К.Кольцов, предложивший своему последователю, В.В. Сахарову, продолжить исследования в данной области.
В.В. Сахаров, работая по замыслу Кольцова, использовал 10% раствор йода. По анализу видимых мутаций, сцепленных с полом, в потомстве второго поколения он нашел передаваемые по наследству новые мутации Notch и "prune".
Впервые в мире в 1938 г. В.В. Сахаров сформулировал идею "о специфическом воздействии мутационных факторов", показал различие природы мутаций, возникших спонтанно, и индуцированных физическими и химическими мутагенами [2, c. 22].
Вопрос № 10. Генетический код, его сущность и основные свойства (триплетность, неперекрываемость, универсальность)
Важнейшим достижением биологии XX в. стало выяснение генетического кода - установление соответствия между последовательностью нуклеотидов молекулы ДНК и аминокислотами молекулы белка. В настоящее время генетический код выяснено полностью.
Каждая аминокислота кодируется тремя (размещенными рядом) нуклеотидами молекулы ДНК или соответствующими (комплементарными) нуклеотидами информационной РНК. Эти нуклеотиды составляют триплеты (тройки, кодоны). Четыре разные нуклеотиды молекулы ДНК - А, Ц, Т, Г (или А, Ц, У, Г молекулы РНК) могут образовывать 64 различных триплета (с учетом последовательности размещение). Все эти триплеты (за исключением трех: УАА, УАГ и УГА) соответствуют 20 аминокислотам, которые входят в состав белков. Некоторые аминокислоты, например триптофан (УГГ), метионин (АУГ), кодируются одним триплетом, другие - двумя (фенилаланин - УУУ, УУЦ; цистеин - УГУ, УГЦ), тремя (изолейцин - АУУ, АУЦ, АУА), четырьмя (глицин - ГГУ, ГЩ; ОГА, ГГГ; пролин - ЦЦУ, ЦЦЦ, ЦЦА, ЦЦГ) и шестью (серин - УЦУ, УЦЦ, УЦА, УЦГ, АГУ, АГЦ) триплетами. Как видно из приведенных примеров, в случае кодирования аминокислот несколькими триплетами эти триплеты отличаются лишь третьей буквой. В случае шести триплетов четыре из них отличаются лишь третьей буквой, а два отличаются три азотистые основания в центральном, втором и третьем кругах кодируют одну аминокислоту, которая сокращенно записана во внешнем круге (от этих четырех) полностью, но между собой они отличаются тоже только третьей буквой (см. код аминокислоты серина) [3, c. 44].
Триплеты УАА, УАГ и УГА (на рис. 1 они обозначены аббревиатурой "ТЕР") выполняют функцию "пунктуации" и не несут генетической информации. Они отделяют информативные участки друг от друга, есть стопкодонами. Именно на них прекращается синтез одной полипептидной цепи. Очевидно, стопкодон - это конечная точка функциональной единицы ДНК цистрона. Начинается синтез следующего цепи триплетом АУГ или ГУГ.
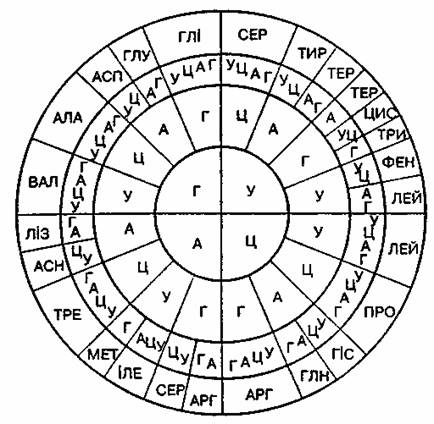
Рис. 1. Генетический код РНК
Генетический код характеризуется вырожденностью (количество аминокислот меньше количества триплетов). Он однозначен (каждый триплет кодирует только одну определенную аминокислоту), универсальный (единый для всех организмов) и не перекрывается [3, c. 45].
Так, на длинной цепи молекулы ДНК закодирована Информация о структуре различных белков. Подобный код является универсальным, поскольку он одинаково функционирует у всех организмов - от вирусов до человека. Участок ДНК, который содержит информацию о первичной структуре конкретного белка, называют структурным геном. Ген может проявляться в нескольких формах - аллелях. Связующим звеном между ДНК ядра и рибосомами, где