Файл: 1. Синтез кетоновых тел Кетоновые тела это способ транспорта ацетильной группы.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 10.01.2024
Просмотров: 262
Скачиваний: 1
Процесс синтеза жирных кислот
Биосинтез жирных кислот наиболее активно происходит в цитозоле клеток печени, кишечника, жировой ткани в состоянии покояили после еды.Условно можно выделить 4 этапа биосинтеза:1. Образование ацетил-SКоА из глюкозы, других моносахаров или кетогенных аминокислот.2. Перенос ацетил-SКоА из митохондрий в цитозоль:· обычно в составе лимонной кислоты, образующейся в первой реакции ЦТК.Поступающий из митохондрий цитрат в цитозоле расщепляется АТФ-цитрат-лиазой до оксалоацетата и ацетил-SКоА.Оксалоацетат в дальнейшем восстанавливается до малата, и последний либо переходит в митохондрии (малат-аспартатный челнок), либо декарбоксилируется в пируват малик-ферментом ("яблочный" фермент).3. Образование малонил-SКоА из ацетил-SКоА.Карбоксилирование ацетил-SКоА катализируется ацетил-SКоА-карбоксилазой, мульферментным комплексом из трех ферментов4. Синтез пальмитиновой кислоты.Осуществляется мультиферментнымкомплексом "синтаза жирных кислот" (синоним пальмитатсинтаза) в состав которого входит 6 ферментов и ацил-переносящий белок (АПБ).Ацил-переносящий белок включает производное пантотеновой кислоты – 6-фосфопантетеин(ФП), имеющий HS-группу, подобно HS-КоА. Один их ферментов комплекса, 3-кетоацил-синтаза, также имеет HS-группу в составе цистеина. Взаимодействие этих групп обусловливает начало и продолжение биосинтеза жирной кислоты, а именно пальмитиновой кислоты. Для реакций синтеза необходим НАДФН. В первых двух реакциях последовательно присоединяются малонил-SКоА к фосфопантетеину ацил-переносящего белка и ацетил-SКоА к цистеину 3-кетоацилсинтазы.3-Кетоацилсинтаза катализирует третью реакцию – перенос ацетильной группы на С2 малонила с отщеплением карбоксильной группы.Далее кетогруппа в реакциях восстановления (3-кетоацил-редуктаза), дегидратации (дегидратаза) и опять восстановления (еноил-редуктаза) превращается в метиленовую с образованием насыщенного ацила, связанного с фосфопантетеином.
Ацилтрансфераза переносит полученный ацил на цистеин 3-кетоацил-синтазы, к фосфопантетеину присоединяется малонил-SКоА и цикл повторяется 7 раз до образования остатка пальмитиновой кислоты. После этого пальмитиновая кислота отщепляется шестым ферментом комплекса тиоэстеразой.
Удлинение цепи жирных кислот
Синтезированная пальмитиновая кислота при необходимости поступает в эндоплазматический ретикулум или в митохондрии. Здесь с участием малонил-S-КоА и НАДФН цепь удлиняется до С18 или С20.4. Синтез триглицеридов и фосфолипидов
Синтез ФЛ и ТАГ
Клетки большинства тканей, особенно печени и жировой ткани, обладают способностью накапливать ТАГ. Жировая ткань функционально специализируется на хранении и мобилизации ТАГ.Вслед за перевариванием пищи в плазме крови увеличивается концентрация глюкозы, инсулина, липопротеинов, богатых ТАГ. Стимулируется активность липопротеинлипазы для гидролиза ТАГ в составе липопротеинов, но снижается активность жиромобилизующей липазы в жировой ткани. Наряду с этим стимулируется образование ТАГ в жировой ткани. Натощак или при повышенной потребности в энергии во время физической работы, повышении уровня катехоламинов, гормона роста, АКТГ и глюкагона в плазме крови, снижении секреции инсулина эти процессы меняются на противоположные - увеличивается липолиз в жировой ткани и высвобождаются жирные кислоты. Они используются в качестве источника энергии. Глицерол используется для глюконеогенеза. Схема депонирования и расщепления нейтрального жира в адипоцитах: Предшественниками для синтеза ТАГ являются активная форма глицерола -глицерол-3-фосфат и активированные жирные кислоты – ацил-КоА.В печени глицерол-3-фосфат может образовываться или в результате фосфорилирования глицерола, или из глюкозы как промежуточный продукт гликолиза. В жировой ткани единственным источником образования глицерол-3-фосфата является гликолиз. Глицерол + АТФ ® Глицерол–3-фосфат + АДФ (глицеролкиназа) Диоксиацетонфосфат + НАДН+Н+ ® Глицерол-3-фосфат + НАД+ Следующим этапом биосинтеза ТАГ является последовательное взаимодействие глицерол-3-фосфата с 2 молекулами ацил-КоА, в результате которого образуется фосфатидная кислота. Реакцию катализирует глицерофосфат-ацилтрансфераза.Глицерол–3-фосфат + 2 ацил-КоА ® Фосфатидная кислота + 2 КоА Фосфатидная кислота Далее при участии фосфатидат фосфатазы происходит гидролиз фосфатидной кислоты с образованием 1,2-диацилглицерола (1,2-ДАГ): Фосфатидная кислота + Н2О ® 1,2-ДАГ+ Н3РО4 На заключительном этапе к 1,2-ДАГ присоединяется третья молекула ацил-КоА с образованием ТАГ. Реакцию катализирует диацилглицерол-ацилтрансфераза:1,2-ДАГ + ацил-КоА ® ТАГ + КоА
Синтез фосфоглицеридов
Биосинтез фосфатидилэтаноламинов (кефалинов) Этаноламин + АТФ ® Фосфоэтаноламин + АДФ (этаноламинкиназа) Фосфоэтаноламин + ЦТФ® ЦДФ-этаноламин + пирофосфат ЦДФ-этаноламин + 1,2-ДАГ ® Фосфатидилэтаноламин (ФЭА) + ЦМФ Биосинтез фосфатидилхолинов (лецитинов)Первый путьФЭА + 3 S-Аденозилметионин (последовательное присоединение) ®® Фосфатидилхолин + 3 S-Аденозилгомоцистеин (метилтрансфераза)Второй путь
Холин + АТФ ® Фосфохолин + АДФ (холинкиназа) Фосфохолин + ЦТФ® ЦДФ-холин + пирофосфат ЦДФ-холин + 1,2-ДАГ ® Фосфатидилхолин + ЦМФБиосинтез фосфатидилсеринов
Первый путь
ФЭА + Серин ® Фосфатидилсерин + Этаноламин Второй путьФосфатидная кислота + ЦТФ ® ЦДФ-диацилглицерол + пирофосфат ЦДФ-ДАГ + Серин ® Фосфатидилсерин + ЦМФ5. Метаболизм липопротеинов
Метаболизм липопротеинов очень низкой плотности (ЛПОНП), липопротеинов промежуточной плотности (ЛППП) и липопротеинов низкой плотности (ЛПНП) Основной функцией этих липопротеинов является транспорт жирных кислот в составе ТАГ из печени к периферическим тканям, особенно в жировую и мышечную. Для синтеза ЛПОНП в гепатоцитах требуется белок апо В-100 и липиды: эфиры холестерола (ЭХС), ТАГ и фосфолипиды (ФЛ).Триацилглицеролы для ЛПОНП синтезируются путем этерификации жирных кислот. Они поступают в гепатоциты из плазмы крови. Источниками жирных кислот являются жирные кислоты, образовавшиеся в результате гидролиза ТАГ жировой ткани, либо синтезированные в печени жирные кислоты. Образование ЛПОНП регулируется после трансляции за счет контроля синтеза апо В-100.Новосинтезированная частица ЛПОНП содержит одну молекулу белка - апо В-100. Другие белковые компоненты, апо С-II, апо С-III и апо Е, поступают на неё от ЛПВП после того, как ЛПОНП попадают в плазму крови. Они требуются для ускорения метаболизма ЛПОНП.Метаболизм ЛПОНП. На эндотелии сосудистой стенки ТАГ в составе ЛПОНП подвергаются действию фермента липопротеинлипазы (ЛПЛ). Необходимым кофактором для проявления активности ЛПЛ является апо С-II. ЛПЛ образуется в клетках многих тканей, среди которых наибольшее значение имеют жировая ткань, скелетная и сердечная мышцы, молочная железа во время лактации. ЛПЛ катализирует гидролиз ТАГ в составе ХМ и ЛПОНП до жирных кислот, моноацилглицеролов (МАГ), в результате ЛПОНП превращаются в кровотоке в ЛППП. Фермент проявляет низкую активность по отношению к МАГ и ФЛ.В жировой ткани синтез ЛПЛ стимулирует инсулин. Тем самым обеспечивается поступление жирных кислот в адипоциты для синтеза и хранения в виде ТАГ. В мышцах ЛПЛ позволяет использовать жирные кислоты для окисления в периоды между приемами пищи, а инсулин подавляет образование этого фермента.Липопротеины промежуточной плотности (ЛППП). Образование ЛППП происходит из ЛПОНП. Около 75 % ЛППП попадает в печень после связывания апо Е с рецепторами для ЛПНП или рецепторами для апо В/апо Е. В печени они полностью разрушаются. Около25 % ЛППП в кровотоке подвергается действию другого липолитического фермента - печеночной липазы (ПЛ). Этот фермент катализирует дальнейшее расщепление ТАГ в составе ЛППП. В результате ЛППП превращаются в ЛПНП.Липопротеины низкой плотности (ЛПНП). ЛПНП удаляются из кровотока путем взаимодействия с рецепторами для ЛПНП (другое их название апо В/Е рецепторы). Доля этого процесса в удалении всех ЛПНП составляет 75 %. Остальная часть удаляется с помощью «мусорных» рецепторов (скэвенджер (scavenger)-рецепторов), имеющих низкую способность связывания. Этот путь получил образное название «путь уборки мусора». Поступление ЛПНП в клетки и их внутриклеточный катаболизм: После связывания ЛПНП комплекс «рецептор—ЛПНП» переносится в клетку посредством эндоцитоза. Затем он сливается с лизосомами и разрушается. Внутриклеточное высвобождение холестерола, происходящее таким путем, вызывает следующие эффекты: а) снижает синтез ключевого фермента образования своего, клеточного, холестерола — ГОМГ-КоА редуктазы; б)
снижает синтез рецепторов для ЛПНП; в) активирует ацил-холестеролацилтрансферазу (АХАТ), которая катализирует образование из метаболически активной формы неэтерифицированного холестерола его метаболически неактивную форму – эфир холестерола ЭХС.
Схема реакции, катализируемой АХАТ:
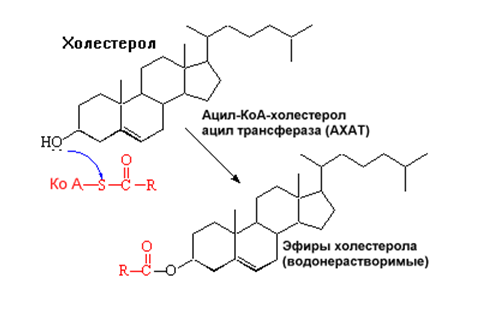
В отличие от регуляторного действия рецепторов к ЛПНП на обмен холестерола в клетках, скэвенджер-рецепторы продолжают транспортировать ХС в клетку без торможения по принципу обратной связи. Тем самым макрофаги превращаются в пенистые клетки. Их образование рассматривается как начальный этап атеросклероза.
Метаболизм липопротеинов высокой плотности (ЛПВП). ЛПВП синтезируются в клетках печени и кишечника. Сразу после секреции ЛПВП имеют вид плоских дисков, содержащих ФЛ. Белковым компонентом их является апо-А. Из тканей и клеточных мембран на них поступает холестерол. Под действием фермента лецитин-холестерол ацилтрансферазы (ЛХАТ) из неэтерифицированного холестерола и жирной кислоты фосфатидилхолина образуются ЭХС. В результате частицы ЛПВП созревают, принимая форму глобулы. Затем ЭХС переносятся на ЛППП, ЛПНП, обломки ХМ с помощью липидтранспортирующего белка (ЛПБ) или апо D.
Жировое перерождение печени
Жировое перерождение печени – это накопление в гепатоцитах триацилглицеролов, которые не окисляются и не выводятся из печени. Причины: действие гепатотоксических ядов (алкоголь, четыреххлористый углерод и другие), хронические заболевания печени, опухоли печени, а также дефицит белков, холина в продуктах питания. При недостатке холина, аминокислот, особенно метионина, снижается синтез лецитинов, но повышается синтез ТАГ, так как у этих соединений есть общий предшественник в процессе синтеза – фосфатидная кислота. Кроме того, при белковом голодании в печени нарушается синтез ЛПОНП, необходимых для выведения синтезированных в ней ТАГ.
Вещества, предупреждающие жировое перерождение печени, называются липотропными факторами. К ним относятся холин, метионин, серин. Кроме того, для предупреждения жирового перерождения печени очень важно наличие в продуктах питания витаминов-синтергистов – В