ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 12.10.2020
Просмотров: 7941
Скачиваний: 25

6
В сенсорных ядрах происходит также и торможение. Тормозные процессы осуществляют
фильтрацию и дифференциацию афферентной информации. Эти процессы обеспечивают контроль
сенсорной информации, который позволяет устранять несущественные, неприятные, избыточные
сигналы.
Н а т р е т ь е м э т а п е
процесс переработки сенсорного сообщения завершается высшим
анализом и синтезом, который происходит в коре большого мозга. После этого уже происходит выбор
или разработка программы ответной реакции организма.
Анализ
заключается в том, что с помощью возникающих ощущений организм различает
действующие раздражители (качественно
–
свет, звук и т.д.) и определяет силу, время и место, т.е.
пространство, на которое действует раздражитель, а также его локализацию (источник звука, света,
запаха).
В центральную нервную систему поступает огромное количество информации от периферических
отделов анализаторов, однако значительная ее часть отсеивается (с помощью механизмов торможения
–
преимущественно
латерального). Эту роль выполняют сенсорные реле
–
промежуточные узловые
структуры сенсорных систем. Они выполняют функции выявления во входных посылках
физиологически важной информации. В результате в сенсорных реле, образующих фильтрующие
(перекодирующие) центры, происходит регулирование суммарного входного информационного потока
в соответствии с требованиями других отделов нервной системы и организма в целом, что обеспечивает
познание внешнего мира и формирование приспособительных реакций.
Синтез
заключается в восприятии предмета, явления и формировании ответной реакции организма.
Восприятие возможно в двух вариантах: когда предмет или явление встречаются повторно или впервые.
Если предмет, явление встречаются повторно, происходит узнавание их в целом по
совокупности
отдельных характеристик раздражителя (гнозис). Узнавание достигается в результате сличения
поступающей в данный момент информации со следами памяти. Без сличения ощущений со следами
памяти узнавание невозможно.
Для осознания действия внешних раздражителей необходимо участие активации специфической
сенсорной системы и активации неспецифической системы
–
ретикулярной формации. Информация о
физических параметрах воспринимаемого объекта передается по специфической сенсорной системе,
активация неспецифической системы определяет уровень бодрствования. Эмоциональная окраска
восприятия зависит от активности лимбической системы, в которую поступают сенсорные сигналы по
нескольким параллельным путям.
Структурно
-
функциональная характеристика коры большого мозга
Кора головного мозга представляет собой многослойную нейронную ткань с множеством складок
общей площадью в обоих полушариях примерно 2200 см
2
, что соответствует квадрату со сторонами 47
х 47 см, ее объем соответствует 40 % массы головного мозга, ее толщина колеблется от 1,3 до 4,5 мм, а
общий объем равен 600 см
3
. В состав коры головного мозга входит 10
9
–10
10
нейронов и множество
глиальных клеток, общее число которых пока неизвестно.
В коре выделяют 6 слоев (
I–VI
), каждый из которых состоит из
пирамидных и звездчатых клеток
(рис. 1). Главная особенность пирамидных клеток заключается в том, что их аксоны выходят из коры и
оканчиваются в других корковых или иных структурах. Название звездчатых клеток также обусловлено
их формой; их аксоны оканчиваются в коре, т.е. речь идет о корковых интернейронах.
В
I – IV
слоях происходит восприятие и обработка поступающих в кору сигналов в виде нервных
импульсов. Покидающие кору эфферентные пути формируются преимущественно в
V–VI
слоях.
Деление коры на различные поля проведено К. Бродманом (1909) на основе цитоархитектонических
признаков
–
формы и расположения нейронов. Ученый выделил 52 поля, многие из которых
характеризуются функциональными и нейрохимическими особенностями.
Согласно гистологическим данным элементарные нейронные цепи расположены перпендикулярно
поверхности коры. Электрофизиологические исследования с погружением микроэлектродов
перпендикулярно поверхности соматосенсорной коры показали, что все встречаемые на пути нейроны
отвечают на раздражитель только одного качества (вида), например на свет. Напротив, при погружении
электрода под углом на его пути попадаются нейроны разной модальности (реагируют на несколько
раздражителей), что свидетельствует о наличии в коре мозга функциональных объединений нейронов,
расположенных в цилиндрике диаметром 0,5
–
1,0 мм. Эти объединения были названы нейронными

7
колонками. Соседние нейронные колонки могут частично перекрываться и осуществлять
саморегуляцию по типу возвратного торможения, а также взаимодействовать друг с другом по
механизму латерального торможения.
Несколько вертикальных колонок объединяются в единую крупную модульную колонку, получая и
перерабатывая большой объем информации. Многократное поступление информации в корковые
модули и ее циркуляция в замкнутых цепях обеспечивают электрические и химические изменения в
нейронах и синапсах, необходимые для организации долговременной памяти.
Рис. 1. Полусхематичное изображение слоев коры головного мозга
(по
K.Brodmann, Vogt
; с изменениями):
а
–
основные типы нервных клеток (окраска по Гольджи);
б
–
тела нейронов (окраска по Нисслю);
в
–
общее
расположение волокон (миелиновых оболочек). Слои пронумерованы от поверхности вглубь. Приведены две
наиболее распространенные системы нумерации. (Объяснение в тексте.)
Отдельный нейрон получает информацию от множества других нейронов и посылает к ним
импульсы. В результате происходит объединение индивидуальных нейронов в системы, обладающие
новыми свойствами, отсутствующими у входящих в их состав единиц. Характерной чертой таких
систем является то, что активность каждого составного элемента в них определяется не только
влияниями, поступающими по прямым афферентным путям каждого элемента, но и состоянием других
элементов системы. Системность в работе нервных образований возникает тогда, когда деятельность
каждой нервной клетки оказывается функцией не только непосредственно поступившего к ней сигнала,
но и функцией тех процессов, которые происходят в остальных клетках нервного центра (П. Г. Костюк).
Обширные связи различных областей мозга между собой делятся условно на
закрепленные
(запрограммированные генетически) и
развивающиеся
в результате сенсорного притока и моторного
опыта. Вследствие этого в мозге помимо жестко закрепленных организуются
распределенные
системы
получения и обработки информации, системы, участвующие в обучении. Любой модуль может входить
в любую систему обработки информации. Распределенные системы мозга анализируют информацию,
поступающую из внешнего мира и внутренней среды организма. Хранится информация с помощью

8
механизмов памяти. Распределенные системы имеют важное значение в формировании потребностей,
мотиваций и эмоций.
По своим
биофизическим свойствам
нейроны коры сходны с другими нервными клетками.
Потенциал покоя
пирамидных клеток составляет от
–
50 до
–
80 мВ, амплитуда их
потенциала действия
–
60 –
100 мВ при длительности 0,5
–
2 мс. Потенциалы действия в нейронах коры возникают в области
аксонного холмика и распространяются не только по аксону, но также по телу и дендритам клетки (по
крайней мере, по проксимальным). Поскольку вслед за потенциалом действия в этих нейронах не
возникает заметной следовой гиперполяризации, частота их импульсации достигает 100 Гц. В нейронах
коры даже у бодрствующих животных частота импульсации, вызванной постсинаптическими
потенциалами, низкая, обычно меньше 10 Гц, а нередко и не достигает 1 Гц. Потенциалы покоя клеток
коры обычно колеблются в диапазоне на 3
–
10 мВ ниже порогового уровня (
Е
кр
), т.е. пороговый
потенциал (
) колеблется и составляет 3
–
10 мВ.
Кортикализация функций
–
возрастание в филогенезе роли коры большого мозга в анализе и
синтезе и в регуляции функций организма, подчинении себе нижележащих отделов ЦНС. Выключение
коры больших полушарий у обезьян и у человека приводит к потере не только локомоции, но и
выпрямительных рефлексов, которые у других млекопитающих сохраняются.
В процессе обработки информации в нейронах коры большого мозга различные отделы коры играют
специфическую роль.
Локализация функций в коре большого мозга
Общая характеристика.
В определенных участках коры большого мозга сосредоточены
преимущественно нейроны, воспринимающие один вид раздражителя: затылочная область
–
свет,
височная доля
–
звук и т. д. Однако после удаления классических проекционных зон (слуховых,
зрительных) условные рефлексы на соответствующие раздражители частично сохраняются. Согласно
теории И. П. Павлова в коре большого мозга имеется «ядро» анализатора (корковый конец) и
«рассеянные» нейроны по всей коре. Современная концепция локализации функций базируется на
принципе
многофункциональности
(но
не
равноценности)
корковых
полей.
Свойство
мультифункциональности позволяет той или иной корковой структуре включаться в обеспечение
различных форм деятельности, реализуя при этом основную, генетически присущую ей, функцию (О.С.
Адрианов). Степень мультифункциональности различных корковых структур неодинакова. В полях
ассоциативной коры она выше. В основе мультифункциональности лежит многоканальность
поступления в кору мозга афферентного возбуждения, перекрытия афферентных возбуждений,
особенно на таламическом и корковом уровнях, модулирующее влияние различных структур, например
неспецифических ядер таламуса, базальных ганглиев на корковые функции, взаимодействие корково
-
подкорковых и межкорковых путей проведения возбуждения. С помощью микроэлектродной техники
удалось зарегистрировать в различных областях коры большого мозга активность специфических
нейронов, отвечающих на стимулы только одного вида раздражителя (только на свет, только на звук и т.
п.), т. е. имеется множественное представительство функций в коре большого мозга.
В настоящее время принято подразделение коры на сенсорные, двигательные и ассоциативные
(неспецифические) зоны (области).
Сенсорные зоны коры.
Сенсорная информация поступает в проекционную кору, корковые отделы
анализаторов (И.П. Павлов). Эти зоны расположены преимущественно в теменной, височной и
затылочной долях. Восходящие пути в сенсорную кору поступают в основном от релейных сенсорных
ядер таламуса.
Первичные сенсорные зоны
–
это зоны сенсорной коры, раздражение или разрушение которых
вызывает четкие и постоянные изменения чувствительности организма (ядра анализаторов по И. П.
Павлову). Они состоят из мономодальных нейронов и формируют ощущения одного качества. В
первичных
сенсорных
зонах
обычно
имеется
четкое
пространственное
(топографическое)
представительство частей тела, их рецепторных полей.
Первичные проекционные зоны коры состоят главным образом из нейронов 4
-
го афферентного слоя,
для которых характерна четкая топическая организация. Значительная часть этих нейронов обладает
высочайшей специфичностью. Так, например, нейроны зрительных областей избирательно реагируют
на определенные признаки зрительных раздражителей: одни
–
на оттенки цвета, другие
–
на
направление движения, третьи
–
на характер линий (край, полоса, наклон линии) и т.п. Однако следует

9
отметить, что в первичные зоны отдельных областей коры включены также нейроны мультимодального
типа, реагирующие на несколько видов раздражителей. Кроме того, там же имеются нейроны, реакция
которых отражает воздействие неспецифических (лимбико
-
ретикулярных, или модулирующих) систем.
Вторичные сенсорные зоны
расположены вокруг первичных сенсорных зон, менее локализованы,
их нейроны отвечают на действие нескольких раздражителей, т.е. они полимодальны.
Локализация сенсорных зон.
Важнейшей сенсорной областью является
теменная доля
постцентральной извилины и соответствующая ей часть парацентральной дольки на медиальной
поверхности полушарий. Эту зону обозначают как
соматосенсорную область
I.
Здесь имеется проекция
кожной чувствительности противоположной стороны тела от тактильных, болевых, температурных
рецепторов, интероцептивной чувствительности и чувствительности опорно
-
двигательного аппарата
–
от мышечных, суставных, сухожильных рецепторов (рис. 2).
Рис. 2. Схема чувствительного и двигательного гомункулусов
(по У. Пенфильду, Т. Расмуссену). Разрез полушарий во фронтальной плоскости:
а
–
проекция общей чувствительности в коре постцентральной извилины;
б
–
проекция двигательной системы
в коре предцентральной
извилины
Кроме соматосенсорной области
I
выделяют
соматосенсорную область
II
меньших размеров,
расположенную на границе пересечения центральной борозды с верхним краем
височной доли,
в
глубине латеральной борозды. Точность локализации частей тела здесь выражена в меньшей степени.
Хорошо изученной первичной проекционной зоной является
слуховая кора
(поля 41, 42), которая
расположена в глубине латеральной борозды (кора поперечных височных извилин Гешля). К
проекционной коре височной доли относится также центр вестибулярного анализатора в верхней и
средней височных извилинах.
В
затылочной доле
расположена
первичная зрительная область
(кора части клиновидной извилины
и язычковой дольки, поле 17). Здесь имеется топическое представительство рецепторов сетчатки.
Каждой точке сетчатки соответствует свой участок зрительной коры, при этом зона желтого пятна
имеет сравнительно большую зону представительства. В связи с неполным перекрестом зрительных
путей в зрительную область каждого полушария проецируются одноименные
половины сетчатки.
Наличие в каждом полушарии проекции сетчатки обоих глаз является основой бинокулярного зрения.
Около поля 17 расположена кора
вторичной зрительной области
(поля 18 и 19). Нейроны этих зон
полимодальны и отвечают не только на световые, но и на тактильные и слуховые раздражители. В
данной зрительной области происходит синтез различных видов чувствительности, возникают более
сложные зрительные образы и их опознание.
Во вторичных зонах ведущими являются 2
-
й и 3
-
й слои нейронов, для которых основная часть
информации об окружающей среде и внутренней среде организма, поступившая в сенсорную кору,
передается для дальнейшей ее обработки в ассоциативную кору, после чего инициируется (в случае
необходимости) поведенческая реакция с обязательным участием двигательной коры.
Двигательные зоны коры.
Выделяют первичную и вторичную моторные зоны.
В
первичной моторной зоне
(прецентральная извилина, поле 4) расположены нейроны,
10
иннервирующие мотонейроны мышц лица, туловища и конечностей. В ней имеется четкая
топографическая проекция мышц тела (см. рис. 2). Основной закономерностью топографического
представительства является то, что регуляция деятельности мышц, обеспечивающих наиболее точные и
разнообразные движения (речь, письмо, мимика), требует участия больших по площади участков
двигательной коры. Раздражение первичной моторной коры вызывает сокращение мышц
противоположной стороны тела (для мышц головы сокращение может быть билатеральное). При
поражении этой корковой зоны утрачивается способность к тонким координированным движениям
конечностями, особенно пальцами рук.
Вторичная моторная зона
(поле 6) расположена как на латеральной поверхности полушарий,
впереди прецентральной извилины (премоторная кора), так и на медиальной поверхности,
соответствующей коре верхней лобной извилины (дополнительная моторная область). Вторичная
двигательная кора в функциональном плане имеет главенствующее значение по отношению к
первичной двигательной коре, осуществляя высшие двигательные функции, связанные с планированием
и координацией произвольных движений. Здесь в наибольшей степени регистрируется медленно
нарастающий отрицательный
потенциал готовности,
возникающий примерно за 1 с до начала
движения. Кора поля 6 получает основную часть импульсации от базальных ганглиев и мозжечка,
участвует в перекодировании информации о плане сложных движений.
Раздражение коры поля 6 вызывает сложные координированные движения, например поворот
головы, глаз и туловища в противоположную сторону, содружественные сокращения сгибателей или
разгибателей на противоположной стороне. В премоторной коре расположены двигательные центры,
связанные с социальными функциями человека: центр письменной речи в заднем отделе средней
лобной извилины (поле 6), центр моторной речи Брока в заднем отделе нижней лобной извилины (поле
44), обеспечивающие речевой праксис, а также музыкальный моторный центр (поле 45),
обеспечивающий тональность речи, способность петь. Нейроны двигательной коры получают
афферентные входы через таламус от мышечных, суставных и кожных рецепторов, от базальных
ганглиев и мозжечка. Основным эфферентным выходом двигательной коры на стволовые и спинальные
моторные центры являются пирамидные клетки
V
слоя. Основные доли коры большого мозга
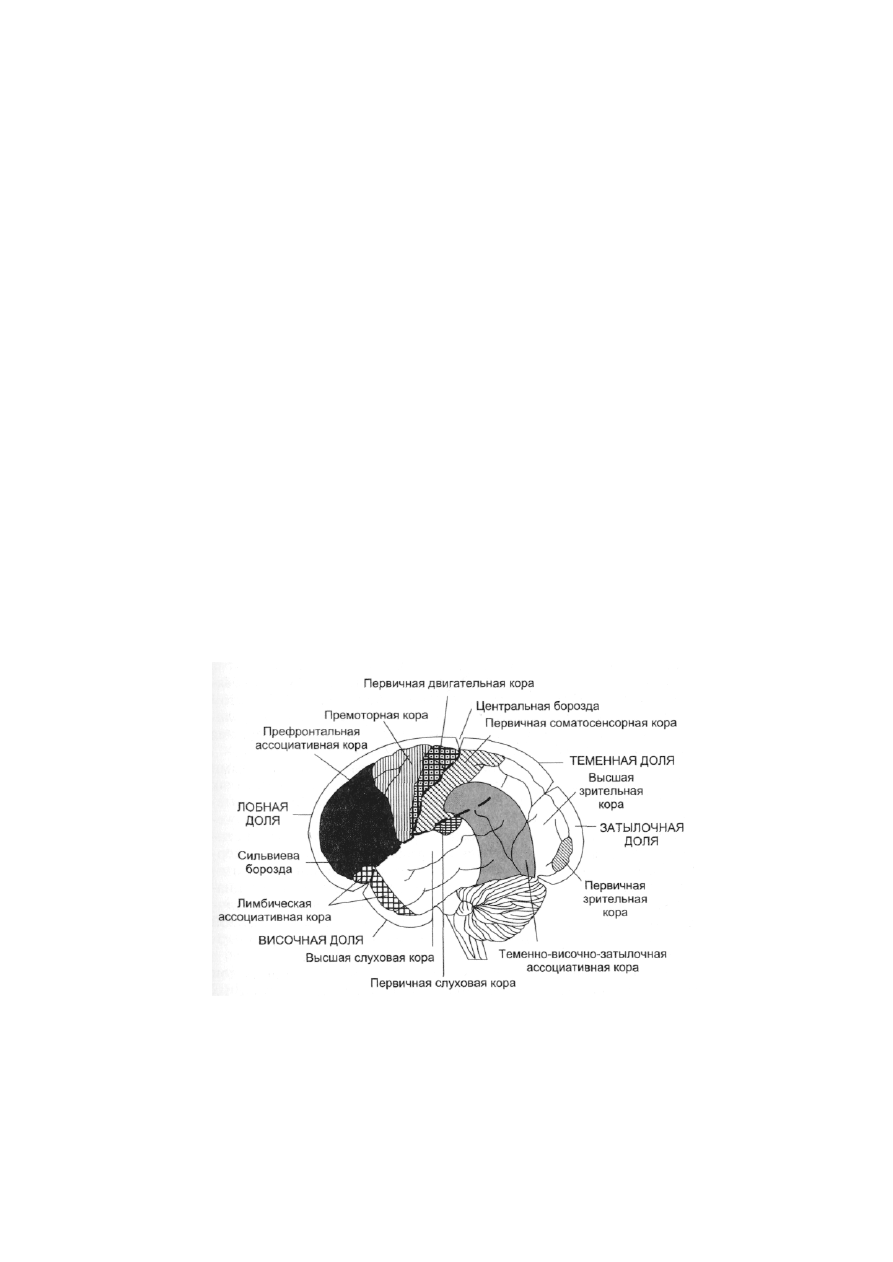
представлены на рис. 3.
Рис. 3. Четыре основные доли
коры головного мозга (лобная, височная,
теменная и затылочная); вид сбоку. В
них расположены первичная
двигательная и сенсорная области, двигательные и сенсорные области
более
высокого порядка (второго, третьего и т.д.) и ассоциативная
(неспецифичная) кора
Ассоциативные области коры
(неспецифическая, межсенсорная, межанализаторная кора)
включают участки новой коры большого мозга, которые расположены вокруг проекционных зон и
рядом с двигательными зонами, но не выполняют непосредственно чувствительных или двигательных