ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 06.11.2023
Просмотров: 179
Скачиваний: 3
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
нуклеиновыми кислотами, углеводами.
Роль нарушений перекисного статуса при заболеваниях пародонта.
Этиопатогенез заболеваний пародонта многогранен, в настоящее время раскрыты его многие ключевые механизмы. Одни авторы считают, что при деструкции пародонта изменения тканей происходят, в основном, за счет микробной инвазии; другие видят причину развития патологического процесса в нарушениях общего и местного иммунитета, некоторые придерживаются мнения о нарушении процессов липопероксидации, и, как следствие, мембранодеструкции. В последние годы накоплен обширный клинический и экспериментальный материал об изменениях местного иммунного, коагулологического потенциала, о сдвигах в системе «ПОЛ – антиоксидантная защита» в генезе патологии пародонта.
Одной из главных причин воспалительных изменений в пародонте является зубная бляшка, способствующая инвазии микрофлоры в ткани и последующему развитию воспалительно-деструктивного процесса с рассасыванием костной ткани альвеолярных отростков челюстей. Влияние иных местных и общих факторов расценивается как второстепенное, не имеющее решающего значения для развития и прогрессирования заболевания. В то же время результаты клинических и экспериментальных работ свидетельствуют о весьма значимой роли нейроэндокринных, нейроиммунных, психологических и других факторов в патогенезе воспалительно-дистрофического процесса в пародонте.
Одним из главных биохимических звеньев начальных патологических изменений в тканях пародонта считается активация свободнорадикального окисления.
Пародонт представляет собой в биохимическом отношении многокомпонентную тканевую систему, находящуюся в морфофункциональном единстве и состоящую из множества структурных элементов – клеток. Биологические мембраны – это надмолекулярные образования, отделяющие клетки друг от друга и осуществляющие компартментализацию внутри них. Они состоят из белков и липидов и представляют собой асимметричные, плоские, замкнутые, бислойные структуры, обладающие внешней и внутренней поверхностями. Для всех мембранных структур присущи реакции перекисного окисления, которые необходимы для самообновления, самоперестройки мембран, их репарации. Определенный уровень скорости реакций свободнорадикального окисления липидов присущ любой нормально функционирующей ткани.
Изменение интенсивности перекисного окисления липидов является ответом клетки на любой стресс при воздействии внешних и внутренних факторов. Он представляет собой процесс непосредственного переноса кислорода на субстрат с образованием пероксидов, кетонов, альдегидов, причем характерной чертой реакции является ее цепной, самоиндуцирующий характер.
На рис. 10 представлена краткая схема реакций перекисного окисления липидов и систем их регуляции.
О2 Fe2+ ФП
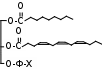

ООС

ОО

InH
RH+In
циклооксигеназа


Fe3+ Цитохром Р450
О2 ОН ОН
О2
НО2



Н2О2
Н2О+О2

+2Н+


СОД
Каталаза

Генерация в дыхательной цепи митохондрий, микросом, функционирование оксидаз и др.
Активные формы
кислорода


Н2О2 НАДФН+Н+




липоксигеназа
ПНЖК
Эйкозанойды
Фосфолипаза А2

Фосфолипиды
Диеновые конъюгаты

КД и СТ

МДА и др. токсические продукты
Гидропероксиды ЖК
InH
RОH+In.
NH2-содержащие соединения (белки, углеводы, нуклеиновые кислоты)
Основания Шиффа


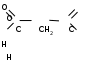

InH - радикальный ингибитор
In. - стабильный радикал антиоксиданта
Рис. 10. Краткая схема процессов ПОЛ и систем их регуляции.
В качестве инициаторов перекисного окисления выступают свободные радикалы, образованные в результате катаболизма и окислительно-восстановительных процессов. В основном это активные формы кислорода, а также оксиды азота и серы. К образованию данных соединений приводят различные полимеры, химизация продуктов сельского хозяйства, определенный уровень радиации и ультрафиолетового излучения, которые способствуют активации перекисного окисления.
Активные формы кислорода образуются не только за счет экзогенных факторов, но и эндогенным путем: в цепи переноса электронов в митохондриях и эндоплазматической сети, в метаболизме пуринов, при активации гексозомонофосфатного шунта в фагоцитирующих клетках и др. Образование свободных радикалов тесно связано с метаболизмом кислорода. В результате ферментативных или спонтанных процессов образуются промежуточные продукты, обладающие повышенной окисляющей способностью: супероксид-анион, синглетный кислород и гидроксильные радикалы. Субстратами для них, в первую очередь в гидрофобной части мембраны, являются полиненасыщенные жирные кислоты. Безусловно, последние – не единственная мишень для атаки свободных радикалов, однако ведущая роль фосфолипидов, содержащих значительное количество полиеновых жирных кислот, в структурно-функциональной организации мембранных образований делает их наиболее вероятными субстратами свободнорадикальных реакций.
Сущность патогенного действия свободных радикалов заключается в их структуре. У свободных радикалов, в отличие от обычных химических веществ, на внешней орбите располагается неспаренный электрон. Такая частица является чрезвычайно реакционноспособной и может атаковать и изменять конформацию молекул, приводя к нарушению их функций. Процесс пероксидации сопровождается: деформацией мембранно-липидного комплекса, увеличением проницаемости для Н+ и воды, ингибированием мембранных ферментов, появлением «пор» в структуре, а в конечном итоге цитолизом и гибелью клетки.
Общими признаками мембранной патологии, вызванной активацией перекисного окисления липидов, являются: увеличение гидрофильности мембран и, вследствие этого, усиление их проницаемости для ионов кальция, других ионов, разобщение тканевого дыхания и окислительного фосфорилирования,
нарушение ферментных функций, ослабление связи фосфолипидов со структурными и рецепторными белками мембран, инактивация тиоловых энзимов, SH-групп аминокислот и белков, повреждение ДНК, набухание и лизис мембран с последующим аутолизом клетки.
Повышение уровня пероксидов липидов и продуктов их дальнейшего катаболизма при ряде заболеваний способствовали созданию представления о реакциях перекисного окисления липидов, как о нежелательном и абсолютно патологическом процессе. Тем не менее, существование ферментативных систем регуляции реакций перекисного окисления свидетельствует о том, что это нормальный метаболический процесс, необходимый для самообновления и самоперестройки мембранных структур, регуляции ионного транспорта и активности мембраносвязанных ферментов, образования многих биологически активных соединений (системных и локальных гормонов).
Ключевым вопросом в понимании механизмов регуляции антиокислителей является их соотношение с уровнем перекисного окисления. Многочисленными исследованиями доказано, что баланс в системе «перекисное окисление – антиоксиданты» является физиологической константой. Более того, показано, что чувствительность этой регуляторной формации сохраняется и при введении экзогенных радикальных ингибиторов, при этом состав фосфолипидов, их окисляемость, скорость расхода антиоксидантов и их концентрация строго уравновешены. Воздействия, изменяющие параметры какого-либо звена этой цепи, приводят к изменениям всей системы, в результате чего свободнорадикальные процессы возвращаются в норме на стационарный уровень. В физиологических условиях продукты ПОЛ инактивируются антиоксидантной системой, которая представлена факторами ферментативной (супероксиддисмутаза, каталаза, система «глутатионпероксидаза-глутатионредуктаза») и неферментативной (витамины А, Е, С, глутатион, цистеин и др.) природы, при недостатке которой воспалительно-деструктивный процесс прогрессирует.
Как повысить антиоксидантный статус организма, нормализовать ослабленные функции его иммунной системы и мобилизовать имеющиеся резервы, показали результаты исследований, проведенных в Научном центре экспертизы и государственного контроля лекарственных средств при Министерстве здравоохранения России. По разработанной в центре методике были выявлены связи между величиной антиоксидантной активности пищевых продуктов, фитопрепаратов и содержанием в них биологически активных веществ в пересчете на флавоноид – кверцетин, присутствующий во многих лекарственных растениях и принятый в качестве стандарта.
Роль нарушений перекисного статуса при заболеваниях пародонта.
Этиопатогенез заболеваний пародонта многогранен, в настоящее время раскрыты его многие ключевые механизмы. Одни авторы считают, что при деструкции пародонта изменения тканей происходят, в основном, за счет микробной инвазии; другие видят причину развития патологического процесса в нарушениях общего и местного иммунитета, некоторые придерживаются мнения о нарушении процессов липопероксидации, и, как следствие, мембранодеструкции. В последние годы накоплен обширный клинический и экспериментальный материал об изменениях местного иммунного, коагулологического потенциала, о сдвигах в системе «ПОЛ – антиоксидантная защита» в генезе патологии пародонта.
Одной из главных причин воспалительных изменений в пародонте является зубная бляшка, способствующая инвазии микрофлоры в ткани и последующему развитию воспалительно-деструктивного процесса с рассасыванием костной ткани альвеолярных отростков челюстей. Влияние иных местных и общих факторов расценивается как второстепенное, не имеющее решающего значения для развития и прогрессирования заболевания. В то же время результаты клинических и экспериментальных работ свидетельствуют о весьма значимой роли нейроэндокринных, нейроиммунных, психологических и других факторов в патогенезе воспалительно-дистрофического процесса в пародонте.
Одним из главных биохимических звеньев начальных патологических изменений в тканях пародонта считается активация свободнорадикального окисления.
Пародонт представляет собой в биохимическом отношении многокомпонентную тканевую систему, находящуюся в морфофункциональном единстве и состоящую из множества структурных элементов – клеток. Биологические мембраны – это надмолекулярные образования, отделяющие клетки друг от друга и осуществляющие компартментализацию внутри них. Они состоят из белков и липидов и представляют собой асимметричные, плоские, замкнутые, бислойные структуры, обладающие внешней и внутренней поверхностями. Для всех мембранных структур присущи реакции перекисного окисления, которые необходимы для самообновления, самоперестройки мембран, их репарации. Определенный уровень скорости реакций свободнорадикального окисления липидов присущ любой нормально функционирующей ткани.
Изменение интенсивности перекисного окисления липидов является ответом клетки на любой стресс при воздействии внешних и внутренних факторов. Он представляет собой процесс непосредственного переноса кислорода на субстрат с образованием пероксидов, кетонов, альдегидов, причем характерной чертой реакции является ее цепной, самоиндуцирующий характер.
На рис. 10 представлена краткая схема реакций перекисного окисления липидов и систем их регуляции.
О2 Fe2+ ФП
ООС
ОО
InH
RH+In
циклооксигеназа
Fe3+ Цитохром Р450
О2 ОН ОН
О2
НО2
Н2О2
Н2О+О2
+2Н+
СОД
Каталаза
Генерация в дыхательной цепи митохондрий, микросом, функционирование оксидаз и др.
Активные формы
кислорода
Н2О2 НАДФН+Н+
липоксигеназа
ПНЖК
Эйкозанойды
Фосфолипаза А2
Фосфолипиды
Диеновые конъюгаты
КД и СТ
МДА и др. токсические продукты
Гидропероксиды ЖК
InH
RОH+In.
NH2-содержащие соединения (белки, углеводы, нуклеиновые кислоты)
Основания Шиффа
InH - радикальный ингибитор
In. - стабильный радикал антиоксиданта
Рис. 10. Краткая схема процессов ПОЛ и систем их регуляции.
В качестве инициаторов перекисного окисления выступают свободные радикалы, образованные в результате катаболизма и окислительно-восстановительных процессов. В основном это активные формы кислорода, а также оксиды азота и серы. К образованию данных соединений приводят различные полимеры, химизация продуктов сельского хозяйства, определенный уровень радиации и ультрафиолетового излучения, которые способствуют активации перекисного окисления.
Активные формы кислорода образуются не только за счет экзогенных факторов, но и эндогенным путем: в цепи переноса электронов в митохондриях и эндоплазматической сети, в метаболизме пуринов, при активации гексозомонофосфатного шунта в фагоцитирующих клетках и др. Образование свободных радикалов тесно связано с метаболизмом кислорода. В результате ферментативных или спонтанных процессов образуются промежуточные продукты, обладающие повышенной окисляющей способностью: супероксид-анион, синглетный кислород и гидроксильные радикалы. Субстратами для них, в первую очередь в гидрофобной части мембраны, являются полиненасыщенные жирные кислоты. Безусловно, последние – не единственная мишень для атаки свободных радикалов, однако ведущая роль фосфолипидов, содержащих значительное количество полиеновых жирных кислот, в структурно-функциональной организации мембранных образований делает их наиболее вероятными субстратами свободнорадикальных реакций.
Сущность патогенного действия свободных радикалов заключается в их структуре. У свободных радикалов, в отличие от обычных химических веществ, на внешней орбите располагается неспаренный электрон. Такая частица является чрезвычайно реакционноспособной и может атаковать и изменять конформацию молекул, приводя к нарушению их функций. Процесс пероксидации сопровождается: деформацией мембранно-липидного комплекса, увеличением проницаемости для Н+ и воды, ингибированием мембранных ферментов, появлением «пор» в структуре, а в конечном итоге цитолизом и гибелью клетки.
Общими признаками мембранной патологии, вызванной активацией перекисного окисления липидов, являются: увеличение гидрофильности мембран и, вследствие этого, усиление их проницаемости для ионов кальция, других ионов, разобщение тканевого дыхания и окислительного фосфорилирования,
нарушение ферментных функций, ослабление связи фосфолипидов со структурными и рецепторными белками мембран, инактивация тиоловых энзимов, SH-групп аминокислот и белков, повреждение ДНК, набухание и лизис мембран с последующим аутолизом клетки.
Повышение уровня пероксидов липидов и продуктов их дальнейшего катаболизма при ряде заболеваний способствовали созданию представления о реакциях перекисного окисления липидов, как о нежелательном и абсолютно патологическом процессе. Тем не менее, существование ферментативных систем регуляции реакций перекисного окисления свидетельствует о том, что это нормальный метаболический процесс, необходимый для самообновления и самоперестройки мембранных структур, регуляции ионного транспорта и активности мембраносвязанных ферментов, образования многих биологически активных соединений (системных и локальных гормонов).
Ключевым вопросом в понимании механизмов регуляции антиокислителей является их соотношение с уровнем перекисного окисления. Многочисленными исследованиями доказано, что баланс в системе «перекисное окисление – антиоксиданты» является физиологической константой. Более того, показано, что чувствительность этой регуляторной формации сохраняется и при введении экзогенных радикальных ингибиторов, при этом состав фосфолипидов, их окисляемость, скорость расхода антиоксидантов и их концентрация строго уравновешены. Воздействия, изменяющие параметры какого-либо звена этой цепи, приводят к изменениям всей системы, в результате чего свободнорадикальные процессы возвращаются в норме на стационарный уровень. В физиологических условиях продукты ПОЛ инактивируются антиоксидантной системой, которая представлена факторами ферментативной (супероксиддисмутаза, каталаза, система «глутатионпероксидаза-глутатионредуктаза») и неферментативной (витамины А, Е, С, глутатион, цистеин и др.) природы, при недостатке которой воспалительно-деструктивный процесс прогрессирует.
Как повысить антиоксидантный статус организма, нормализовать ослабленные функции его иммунной системы и мобилизовать имеющиеся резервы, показали результаты исследований, проведенных в Научном центре экспертизы и государственного контроля лекарственных средств при Министерстве здравоохранения России. По разработанной в центре методике были выявлены связи между величиной антиоксидантной активности пищевых продуктов, фитопрепаратов и содержанием в них биологически активных веществ в пересчете на флавоноид – кверцетин, присутствующий во многих лекарственных растениях и принятый в качестве стандарта.