ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 18.11.2021
Просмотров: 5583
Скачиваний: 10
291
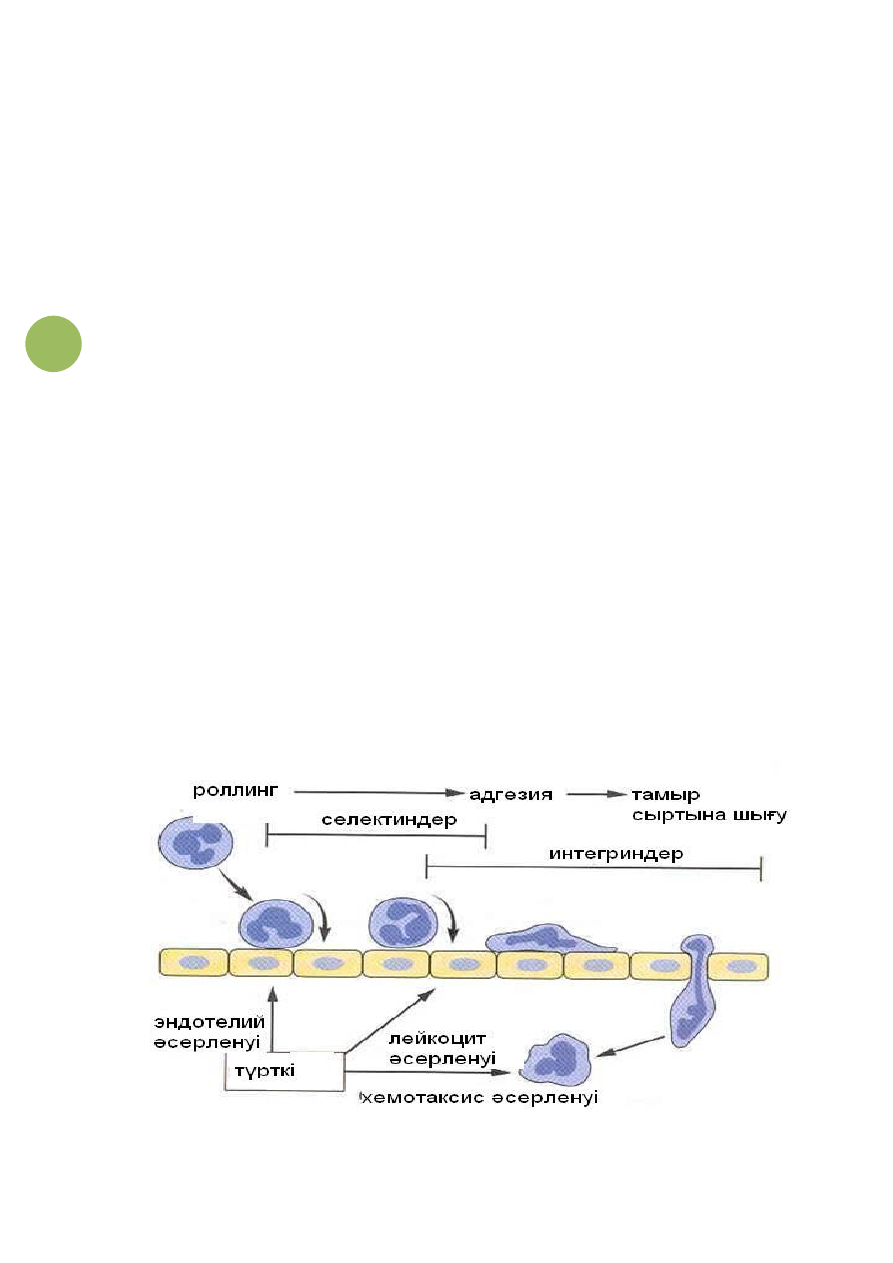
7-сурет.
Лейкоциттердің
эндотелий
жасушаларының
беттерінде тербеліп тұруы.
Артынан
қабыну
медиаторларының
әсерлерінен
лейкоциттердің интегриндері әсерленіп, олардың жәрдемімен
эндотелий жасушаларының иммундық глобулиндерге ұқсас
адгезиялық молекулаларымен қатты жабысады. Содан
лейкоциттер тамырдың ішкі қабығына жармасып алады да,
өздерінің жалған аяқшаларын эндотелий аралық саңылау
арқылы тамыр сыртына шығарады. Артынан олар белсенді
қозғалыстар жасап, іштеріндегі құрылымдарын тамыр сыртына
өткізеді (8-сурет).
Бұл кезде эндотелий жасушалары лейкоциттерден шығатын
медиаторлардың әсерлерінен жиырылады. Содан эндотелий
жасушалары дөңгеленіп, арасындағы саңылаулары кеңиді.
Лейкоциттер шығып кеткен соң эндотелий жасушаларының
жанасуы қалпына келеді.
Эндотелий қабатынан өткен соң лейкоцитке тіректік
мембрана арқылы өтуі қажет. Бұл мембрананың қалыңдығы
40-60 нм және ол гликозамингликандары көп біркелкі зат
пен коллаген талшықтарынан тұрады. Әртүрлі ядролы
лейкоциттер тіректік мембрана арқылы өту кезінде оған
өздерінің
ферменттерімен
(эластаза,
коллагеназа,
гиалуронидаза) әсер етеді. Олар көрсетілген мембрананың
молекулалық
құрылымын
өзгертіп,
оның
өткізгіштігін
жоғарылатады. Бұл үрдісте ферменттерден басқа катиондық
нәруыздарда белгілі қызмет атқарады.
8-сурет. Лейкоциттердің тамыр сыртына шығу кезеңдері.

292
Лейкоциттердің тамыр сыртына шыққан соң қабыну
ошағында қозғалуы байқалады. Бұл хемотаксис құбылысымен
түсіндіріледі.
Хемотаксис (лат. chеmіа -химия; грек.
tахіs - тартылып қозғалу) химиялық қоздырғыштардың
әсерлерінен лейкоциттердің осы қоздырғыштардың көп жағына
қарай бағытталған қозғалысы.
Хемотаксис лейкоциттердің
тамыр сыртына шығуының барлық сатыларында, әсіресе тамыр
сыртында және тамырлары жоқ тіндердегі (көздің сірі
қабығында) олардың қозғалыстарында маңызды орын алады.
Хемотаксис
туындататын
заттарды
хемоаттрактанттар
(химиялық тартқыштар) дейді. Олар белгілі бір лейкоциттің
түріне ғана әсер ететін –
спецификалық
және олардың
барлық түрлеріне бірдей әсер ететін -
бейспецификалық
болып екі түрге ажыратылады.
Спецификалық химиялық тартқыштарға:
●
нейтрофилдердің
ғана
хемотаксисін
туындататын
интерлейкин-8-ді;
● моноциттердің хемотаксисін туындататын ЭМ-Си-Пи-1 (
ағылш. МСР-1, monocyte chemotactic protein-1)–ді;
● лимфоциттерді тартатын лимфотаксинді;
● эозинофилдерді тартатын эотаксинді – келтіруге
болады.
Бейспецификалық
химиялық
тартқыштарға
мыналар
жатады:
●
микроорганизмдер
және
олардың
өнімдері
(липополиқанттар, пептидтер);
● комплемент жүйесінің құрамбөлшектері (С
5а
), Хагеман
факторы, калликреин, брадикинин;
● жасушалардың зат алмасу және бүліну өнімдері (цАМФ,
коллагеннің ыдырау өнімдері);
● лейкотриендер В
4
, А
4
тромбоксандар В
2
, А
2
және
иммундық кешендер.
Көрсетілген заттар лейкоциттердің мембранасындағы
химиялық әсерлерді сезетін рецепторларды қоздырады.
Осыдан лейкоциттердің қабықтарында иондардың тасымалдануы
және
мембраналық
потенциал
өзгереді.
Олардың
цитоплазмасында Са
2+
- иондарының мөлшері артады, содан
жасуша
ішілік
майда
талшықтардың
жиырылуы
болып,
лейкоциттердің
сыртқы
пішіні
өзгереді
де,
жалған
аяқшалары
пайда
болады.
Лейкоциттердің
іштеріндегі
түйіршіктері сыртқы қабығына қарай ауысады.
Лейкоциттердің
қабыну
ошағына
шығуында
белгілі
заңдылық
байқалады.
Мәселен,
алдымен
нейтрофилдік
лейкоциттер, содан кейін моноциттер, соңынан лимфоциттер
шығады. Қабыну ошағында нейтрофилдер алғашқы 2-4 сағатта,
293
ал бір ядролы лейкоциттердің саны 18-24 сағаттан кейін
жоғары дәрежеге жетеді. Соңғылардың ішінде алдымен
моноциттер, содан кейін лимфоциттер көбейеді. Бұндай
заңдылық әртүрлі лейкоциттердің беттеріндегі хемотаксис
туындататын заттарды қабылдайтын рецепторлардың әртүрлі
болуымен түсіндіріледі. Сондықтан әртүрлі лейкоциттер
әртүрлі хемотаксис туындататын заттарға жауап қайтарады.
Экссудаттың жасушалық құрамы қабынудың пайда болу
себебімен үлкен дәрежеде байланысты. Егер қабыну ірің
туындататын микробтардың (стафилококктар, стрептококктар)
әсерінен дамыса, онда экссудатта нейтрофилдік лейкоциттер
басым болады, ал қабыну иммундық негізде (аллергия) болса
немесе қарапайым жәндіктермен, құрттармен шақырылған
болса, онда эозинофилдер көп болады. Созылмалы қабыну
кезінде (туберкулез, мерез т.б.) қабынулық жалқықта бір
ядролы жасушалар (лимфоциттер, моноциттер) басым болады.
Қабыну ошағында лейкоциттердің негізгі қызметі болып,
бөтен бөлшектерді жұтып жеп қою (фагоцитоз) есептеледі.
Фагоцитоз
кезінде
жемір
лейкоциттер
күрделі
қорғаныстық бейімделу тетіктерін қосады. Олар желінетін
бөлшектерге бактерияларды жоятын әсер етуден басқа,
біршама
қабыну
медиаторларын
(простагландиндер
мен
лейкотриендер
т.б.)
шығарады,
тотығу-тотықсыздану
реакцияларын
шұғыл
көтереді,
иммундық
жауапқа
жасушалардың бірігуін дайындайды.
И.
И.
Мечников
барлық
фагоциттерді
микро-және
макрофагтар
деп
бөлді.
Біріншісіне
нейтрофилдік
гранулоциттер
жатады.
Олардың
цитоплазмасында
лизосомалардан тұратын азурофилдік түйіршіктер және
лизосомалық
емес
ферменттері
(лизоцим,
негіздік
фосфатаза, лактоферрин т. б.) катиондық нөруыздар мен
нейтрофилдік пироген (дене қызуын көтеретін фактор)
болатын ұсақ арнайы (спецификалық) түйіршіктер бар.
Сонымен қатар оларда сөлденістік көпіршіктер болады.
Бұлар нейтрофилдердің тамыр сыртына шығуын және олардың
қабыну ошағында белсенді түрде қозғалып жүруін қамтамасыз
етеді. Азурофилдік және арнайы түйіршіктердің ішкі
мембраналарында Н
+
- насосы болады. Ол сутегі иондарын
фагоциттің цитоплазмасынан фагосоманың ішіне ауыстырады.
Осыдан оның ішінде рН 4-5-ке дейін төмендейді де, ондағы
микробтар тіршілігін жояды. Артынан бұл тіршілігін жойған
микробтар фагосоманың ішінде азурофилдік түйіршіктердің
қышқыл гидролазаларының қатысуымен ыдыратылады.
Бір
ядролы
фагоциттер
сүйек
кемігінде
промоноциттерден өніп, қанға моноциттер түрінде түседі.
Ары қарай қанның моноциттері тіндік макрофагтарға
294
айналады. Оларга гистиоциттер, лимфоидты тіннің, өкпе
ұяшықтарының, іш пердесінің, өкпеқаптың макрофагтары,
Купфер жасушалары, остеокластар, микроглия т.с.с. жатады.
Макрофагтар, нейтрофилдерге қарағанда көптеген белсенді
заттар өндіреді. Оларда:
● түзілістік үрдістерді арттыратын заттар;
● интерлейкин - 1 (эндогендік пироген, Т-хелпер
жасушаларының, лимфоциттердің эмиграциясын, фибробластың
пролиферациясы
мен
қызметін
арттырады,
простагландиндердің түзілуін жігерлендіреді);
●
интерферон
(вирустардың
өсіп-өнуін
тежейді,
нейтрофилдерді әсерлеп, фагоцитозды күшейтеді);
● жіті қабынудың сатыларына тән нәруыздардың (α-
гликопротеид, α-антитрипсин, С-реактивті нәруыз т.с.с.)
түзілуін арттыратын заттар өндіріледі.
Көптеген
қоздырғыштар
(вирустар,
бруцелл
микобактериялары, сальмонелдер т. б.) тек макрофагтармен
фагоциттелінеді. Макрофагтар жасушаларды (өзгерген меншік
жасушаларды, ауыстырылып қондырылған тін немесе өспе
жасушаларын) ыдыратып жібере алады. Бұл кезде макрофагтар
жасушаларға
лизосомалық
ферменттер,
комплемент
компоненттері, тотықтырғыштар және қабыну медиаторларымен
әсер етеді.
Фагоцитоз төрт сатыда өтеді: 1. жақындасу, 2. жабысу,
3. жұту, 4. қорыту. Жемір жасушалардың желінетін
бөлшектерге жақындасу жолында хемотаксистің маңызы өте
үлкен (жоғарыда келтірілді). Кейде желінетін затқа
фагоциттердің
жабысуы
олардың
сыртқы
беттерінде
микробтардың мембранасына кіретін немесе тіршілігін
жойған меншік жасушалардың сыртқы беттерінде болатын
молекулаларға рецепторлардың болуымен байланысты болады.
Ал, көпшілік жағдайларда организмге енген микробтарға
фагоциттердің жабысуы опсониндердің қатысуымен дамиды.
Опсониндер қабыну ошағына өткен қан сұйығында болады.
Оларға иммундық глобулиндер G және М, комплементтің С
3в
құрамбөлшегі, С-реактивті нәруыз, лизоцимді жатқызуға
болады. Опсониндер бір жағымен желінетін затқа, екінші
жағымен лейкоциттің сыртқы қабығына жабысып, оларды
бірімен бірін байланыстырады. Мәселен, IgG өзінің Fab
бөлшегімен микробтың бетіндегі антигенмен, Fc-бөлшегімен
лейкоциттің сыртқы бетіндегі рецептормен байланысады.
Осылай опсониндердің әрі микробпен, әрі лейкоцитпен
байланысуын опсонизация дейді.
Желінетін зат фагоцитке жабысқаннан кейін ары қарай
оның жұтылуы болады. Бұл кезде фагоцит өзінің жалған
аяқшаларын шығарып, желінетін затты қоршап алады. Осының
295
нәтижесінде
фагоциттің
ішінде
айналасынан
қабықпен
қоршалған, фагосома – деп аталатын, қалта пайда болады.
Оған
фагоциттің
ішіндегі
азурофилдік
түйіршіктері
(лизосомалары) жақын келеді де, фагосома мен лизосоманың
сыртқы қабықтары бірігіп кетеді. Осылай фаголизосома
қалыптасады. Ары қарай жұтылған зат, фагоциттің ішкі
ортасынан
оқшауланған,
фаголизосоманың
ішінде
ыдыратылады. Бұл фагоцитоздың жасуша ішілік қорытылу
сатысы. Егер желінетін зат тірі микроб болатын болса,
онда ол фаголизосоманың ішінде тіршілігін жояды.
Фагоциттерде екі түрлі жолмен бактерияны жоятын әсер
етіледі. Біріншісі
оттегіге тәуелді
және
екінші жолы
оттегіге тәуелсіз
болады.
Оттегіге тәуелді бактерия жоятын әсер
оттегінің
белсенді бос радикалдарының құрылуы арқылы іске асады.
Бұл радикалдар фагоциттердің бактерияға жабысуынан кейін
өндіріле бастайды. Әдетте фагоциттер қызмет атқару үшін
анаэробтық
гликолиз
жолымен
түзілген
энергияны
пайдаланады. Ал, фагоциттер желінетін затпен жабысқаннан
кейін оттегіні тым артық пайдалана бастайды. Осындай
құбылысты «
тыныстық жарылыс»
деп атайды. Оның дамуы НАДФН
- оксидаза ферментінің белсенділігі артуымен байланысты
болады. Осыдан оттегінің молекуласы НАДФН молекуласынан
бір электронды қосып алады да, супероксиданион радикалға
айналады. Бұл кезде глюкозаның гексозомонофосфаттық
жолмен тотығуы артады да, НАДФН-тың кемістігі толтырылып
тұрады.
Супероксиданион
радикалдың
біршамасы
супероксиддисмутаза
ферментінің
қатысуымен
сутегінің
асқын тотығына (Н
2
О
2
) айналады. Ол валенттігі өзгеріп
тұратын металдардың (Fe, Cu) қатысуымен супероксид
радикалымен
әрекеттесіп,
өте
белсенді
гидроксил
радикалына (ОН
•
) айналады. Бұлар фагосоманың ішінде
құрылады және бактериялардың қабықтарын бүлдіретін әсер
етеді. Осы жолмен фагоциттердің барлық түрлері желінген
бактерияларды жояды.
Оттегіге тәуелсіз бактерия жоятын әсер
нейтрофилдер
мен
эозинофилдерде
болады.
Нейтрофилдердің
лизосомаларында
миелопероксидаза,
эозинофилдерде
пероксидаза ферменттері өте күшті бактерия жойғыш әсер
етеді. Бұл ферменттер моноциттер мен макрофагтарда
болмайды.
Миелопероксидаза (МПО) нейтрофилдердің азурофилдік
түйіршіктерінде болатын фермент. Ол галоген иондарымен
(жиі хлор ионымен) сутегінің асқын тотығының әрекетесуін
қамтамасыз етеді де, гипохлорид анионы (OCl
‾
) түзілуіне