ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 18.11.2021
Просмотров: 5595
Скачиваний: 10

316
Керісінше, жасуша ішінде осмостық қысым сыртындағыдан
төмен болғанда жасушаның бүрісіп қалуы байқалады. Содан
жасушаның құрылымы мен атқаратын қызметтері бұзылады (Су
мен электролиттер алмасуының бұзылыстарын қараңыз).
ИММУНДЫҚ (АЛЛЕРГИЯЛЫҚ) ӘСЕРЛЕР.
Антиденелер мен сезімталдығы көтерілген лимфоциттер,
комплементтің құрамбөлшектері, цитокиндер, табиғи жендет
жасушалар, макрофагтар жасуша мембраналарына бүліндіргіш
әсер етеді. Макрофагтар бүлінген жасушалардың бөлшектерін
жеп қояды. Фагоцитоздық қызметтің негізінде лейкоциттерде
еркін
радикалды
тотығудың
артуы,
оттегінің
бос
радикалдары өндірілуінің көбеюі маңызды орын алады.
Сонымен бірге, нысана-жасушалардың бүліністері антиген-
антидене кешендерінің, комплемент құрамбөлшектерінің ( С
3а,
С
5в
, С
8
, С
9
т.б.), цитотоксиндік Т-киллер және табиғи
жендет (киллер) жасушалардың, өспені жоятын фактордың,
гамма-интерферонның т.с.с. цитокиндердің, лизосомалық
ферменттердің, оттегінің бос радикалдарының әсерлерінен
дамиды (Аллергия тақырыбын қараңыз).
Биологиялық мембраналардың қандайда болмасын жолмен
бүлінуі
олардың
барлық
қызметтерінің
бұзылыстарына
әкеледі.
Мембрананың тосқауылдық қызметінің бұзылыстары.
Қанықпаған май қышқылдары асқын тотығып кетуден
мембрананың тұтқырлығы көтеріледі, қимылдық қозғалыстары
азаяды. Асқын тотығулық «кластерлер» деп аталатын,
мембранада тесіктер пайда болады. Осыдан мембрананың
өткізгіштігі көтеріледі, жасуша ішінде және сыртындағы
иондардың қалыпты арақатынасы өзгереді, иондық дисбаланс
дамиды. Жасуша ішінен калий иондары сыртқа шығып, натрий
иондары ішіне түседі. Натрий иондарымен бірге су енеді,
сөйтіп жасуша ішінде ісіну пайда болады. Бұлшықет
жасушаларының ішіне көптеп Са
2+
иондары кіруден олардың
сіресіп қалуына әкелетін қатты жиырылулары болады. Жасуша
іші мен сыртында иондардың арақатынасы бұзылудан қозымды
тіндерде
мембраналық
потенциал,
әрекеттік
потенциал
бұзылады. Осыдан миокард жасушалары бүлінгенде ЭКГ-де, ми
нейрондары
бүлінгенде
ЭЭГ-да,
бұлшықет
жасушалары
бүлінгенде электромиографияда (ЭМГ) өзгерістер пайда
болады.
Сонымен бірге Са
2+
иондары кәлмодулинмен байланысып,
фосфолипаза, қышқыл протеаза ферменттерін әсерлейді.
Осылардың нәтижесінде кері айналып соғу шеңбері бойынша
жасуша мембранасының қызметтері бұзылуын одан сайын
күшейтеді. Мембрана өткізгіштігі көтерілуіне байланысты
жасуша ішіндегі ферменттер сыртына шығып кетеді, жасуша
317
аралық сұйықтармен қанға түседі. Мәселен, бауыр, жүрек
жасушалары мембраналарының өткізгіштігі жоғарылауынан
қанда
аспартатаминотрансфераза,
алатаминотрансфераза,
лактатдегидрогеназа,
креатинфосфокиназа
т.
б.
ферменттердің деңгейлері көтеріледі. Бұлшықет жасушалары
мембраналарының өткізгіштігі көтерілуінен қанда миоглобин
көбейеді. Бұлардың қандағы деңгейін анықтау клиникада
дертті аңғару үшін кеңінен қолданылады.
Мембраналардың
рецепторлық
қызметтерінің
бұзылуы.
Мембраналарда көптеген рецепторлар (қабылдағыштар) болады.
Бұларға әсер ететін заттарды лигандалар деп атайды. Оларға
нейромедиаторлар,
цитокиндер,
пептидтік
гормондар,
антигендер, стероидтық және тиреоидтық гормондар т.с.с.
заттар
жатады.Лигандалардың
әсерлерінен
рецепторлар
қоздырылып, екінші қатардағы дәнекерлер өндіріледі де
жасуша қоздырғышқа жауап қайтарады.
Мембраналық рецепторлар жасушаға әсер еткен дабылды
қабылдап, оны жасуша ішіне қабарлайды. Осыдан жасуша
ішіндегі химиялық үрдістерді реттеуге қатысатын екіншілік
дәнекерлер өндіріліп, жасушаның қызметтері атқарылады. Бұл
екіншілік дәнекерлерге цАМФ, цГМФ, инозитолтрифосфат,
диацилглицерол, Са
2+
-иондары жатады.
Рецептордан жасушаның ішіне берілетін хабардың негізгі
жолының
бірі
болып,
мембраналық
аденилатциклаза
ферментінің әсерленуі есептеледі. Осының нөтижесінде АТФ
молекуласынан циклдік АМФ түзіледі. Ол өз алдына
протеинкиназа ферменттерін әсерлеп, фосфорлау үрдістерін
күшейту арқылы көптеген жасуша ішіндегі үрдістерді
сергітеді
(9-сурет).
Осыдан
мембранада
белсенді
тасымалдау, нәруыздардың түзілуі, ферменттердің әсерленуі
күшейеді де, жасушаның биологиялық қызметі көтеріледі.
Мысал ретінде фосфорила-заның әсерлену жолын келтіруге
болады.
Глюкагон
немесе
адреналиннің
әсерлерінен
аденилатциклазаның белсенділігі көтеріліп циклдік АМФ
түзіледі. Ол өз алдына протеинкиназа ферментін әсерлеп,
фосфорилаза В-ны фосфорилаза А-ға айналдырады. Соңғысы
жасушадағы
гликогенді
ыдыратып,
глюкозаның
мөлшерін
көбейтеді. Әрі қарай глюкозаның тотығуы нәтижесінде
тіршілікке қажетті макроэргиялық қосындылар түзіледі. 1
моль глюкоза тотығуынан 40 моль АТФ өндіріледі.
Рецепторлардан жасушаның ішіне берілетін хабардың тағы
бір маңызды жолы болып Са
2+
иондары арқылы өтетін үрдістер
есептеледі (9-сурет).
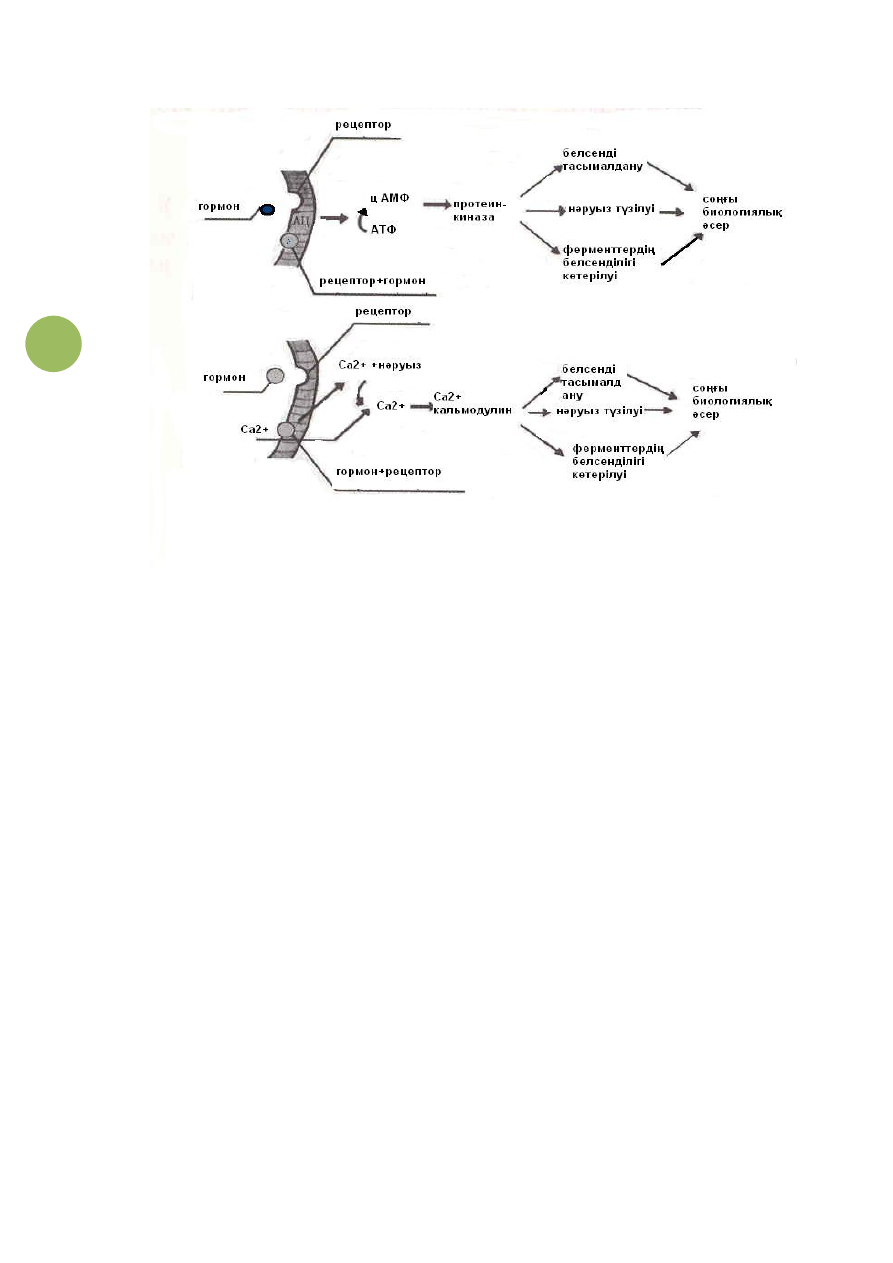
318
9-сурет. Жасуша ішіндегі өтетін үрдістерді реттеу
жолдары.
Мәселен,
гормондар
жасуша
мембранасындағы
рецепторлармен байланысып, Са
2+
иондарының жасуша ішіне
енуін күшейтеді. Осының нәтижесінде жасуша ішінде Са
2+
иондары көбейеді. Са
2+
иондары кәлмодулинмен байланысып
фосфорилазаны әсерлейді. Артынан гликогеннің ыдырауы
артып, глюкозаның деңгейі көтеріледі. Сонымен бірге,
жасуша ішіне хабар жеткізілуіне фосфатидилинозитолдың
өнімдері қатысады. Олар G-нәруыздарымен бірігіп әсер
етеді. Мембранадағы инозитолфосфатаза (фосфолипаза С)
әсерленуінен
фосфатидилинозитол-4,5-дифосфат
инозитолүшфосфатқа және диацилглицеринге ыдырайды. Бұлар
жасуша ішінде Са
2+
- иондарын көбейтіп, протеинкиназа С –
дің белсенділігін көтереді. Осыдан көптеген жасуша ішілік
нәруыздардың фосфорлануы болады, жасуша құрылымына қажетті
нәруыздардың әсерленуі, олардың өсіп-өнуін қадағалайтын
гендік
бағдарламаның
әсерленуі
байқалады,
простагландердің түзілуі артады.
Бұл
көрсетілгендердің
барлығы
цитоплазмалық
мембрананың жағдайына байланысты. Мембрананың қанықпаған
май
қышқылдарының
асқын
тотығуға
ілігіп
кетуінен
микротұтқырлығы көтерілуі рецепторлардың липидтік екі
қабаттың
бойымен
қозғалуын
шектейді.
Мембрананың
рецепторлық қызметтері бұзылуы нәтижесінде жасушалардың
нейрогуморалдық
реттеу
хабарларына
жауабы
өзгереді,
319
хабардың жасуша ішіне берілуі бұзылады. Осыдан жасушада
заттардың алмасулары және оның қызметі өзгереді.
Жасушалардың бірін-бірі тану үрдістерінде және олардың
өзара әсерлерінде мембрана бетінде орналасқан хабаршы
молекулалардың маңызы үлкен. Оларға тіндік үйлесімділік,
топтық ажыратулық, тінге арнайылық т. б. антигендер
жатады. Бұлардың құрылымы мен ерекшеліктерінің өзгеруі
жасуша-аралық
әсерлердің
бұзылуына
және
қажетсіз
жасушалардың
жойылуына
әкеледі.
Мысалы,
бүлінген
жасушалар сыртындағы гликопротеидтерді жоғалтады. Көптеген
ауру туындататын микроорганизмдер (тырысқақ вибрионы,
тұмаудың вирусы т. б.) нейраминидаза ферментін шығарып,
жасуша қабығындағы антигендік қасиеті бар, жасырын
бөлшектерін
ашады.
Бұл
бөлшектер
қалыпты
аутоантиденелермен
байланыса
алады
және
организмнің
иммундық жауабын шақырып, артынан бүлінген жасушаларды
жоятын иммундық әсерлердің (комплемент, макрофагтар)
шабуылына ілінеді. Осылай олар организмнен аластанады.
Мембрана өзіне вирустық нәруыздың бөлшектерін бекітеді.
Осыдан бүлінген жасушаларды жендет жасушалар тез байқап,
жойып отырады.
Жасуша рецепторларының бұзылыстары мына жағдайларда
болуы ықтимал:
● рецепторлардың сезімталдығы өзгеруден;
● рецепторлардың саны өзгеріп кетуден;
●
рецепторлық
нәруыздардың
конформациялық
бұзылыстарынан;
● рецепторлардың айналасын қоршаған липидтердің
өзгерістерінен. Бұл өзгерістердің барлығы мембрананың
бұзылыстарында байқалады.
Екіншілік дәнекерлердің ауытқулары да жасуша ішілік
үрдістердің реттелулерінің бұзылыстарына әкеледі. Мәселен,
жүрек
ет
жасушаларында
цАМФ
жиналып
қалудан,
кардиомиоциттерде
мембраналық
потенциал
қалыптасуы
бұзылып, жүрек аритмиялары дамиды. Бронхиалық демікпе
кезінде кеңірдекшелердің тегіс еттерінде цАМФ азайып
кетуден, олар қатты жиырылып қалады. Саркоплазмалық
торшалардың Са
2+
-АТФ-аза ферментінің белсенділігі
төмендеуден
Са
2+
-иондарының
бұлшықет
жасушаларының
іштерінде жиналып қалуы, олардың сіресіп жиырылуына
әкеледі.
Мембраналардың насостық қызметтерінің бұзылуы.
Жасуша
сыртындағы
мембрананың
Nа
+
-,
К
+
-АТФаза
ферментінің
белсенділігі төмендейді. Сондықтан жасуша ішінде жиналған
натрий иондары сыртқа шығарылып, калий иондарының оның
ішіне
кіруі
бұзылады.
Саркоплазмалық
ретикулум
320
мембранасында
Са
2+
-АТФаза
ферментінің
белсенділігі
төмендейді. Сөйтіп Са
2+
иондары саркоплазмада жиналып
қалады.
Мембранадағы
насостардың
әлсіреуі
май
қышқылдарының
асқын
тотықтарының
(RОН°)
әсерлерінен
болатыны белгілі. Сонымен бірге олардың бұзылыстарында
энергия жеткіліксіздігінің де маңызы үлкен (төменде
келтірілген).
Мембраналардың
каналдық
қызметтерінің
бұзылуы.
Мембранадағы
липидтердің
асқын
тотығуынан
және
фосфолипаза, липаза ферменттерінің артық әсерленулерінен,
фосфолипидтер ыдыратылады. . Са
2+
- тәуелді протезалардың
ықпалдарынан
канал
құратын
нәруыздар
құрылымдық
өзгерістерге ұшырайды. Осыдан және бұл нәруыздардың
молекулаларында амин қышқылдарының кеңістікте орналасу
тәртіптерінің
өзгерістері
нәтижесінде
мембранадағы
каналдардың қызметтері бұзылады
Мембраналардың қызметтерінің бұзылыстары, жасушаның
сыртқы
мембранасында
ғана
болмай,
жасуша
ішіндегі
құрылымдардың мембраналарында да болады. Осыған байланысты
митохондрийлардың мембраналары бұзылудан энергия түзілуі
бүлінеді. Қалыпты жағдайда тотығу-тотықсыздану реакциялары
нәтижесінде босаған энергияның бір бөлшегі жылу ретінде
тарайды, ал қалғаны (60-%-ға жуығы) энергиялық қосындылар
(АТФ, КрФ) түрінде жиналады. Соңғылары артынан тіршілікке
қажетті организмнің әртүрлі қызметтерінде (ет жиырылу,
бездердің сөлденісі, ойлау т. с. с.) пайдаланылады.
Митохондрийлардың мембраналары бұзылыстарынан онда су,
Са
2+
иондары, липофуксин, ферритин т.с.с. заттар артық
жиналып қалады. Сонымен бірге олардағы ферменттер жүйесі
әлсірейді.
Сондықтан
митохондрийларда
тотығу
мен
фосфорланудың ажырауы болады да макроэргиялық қосындылар
аз түзіледі. Осыдан организмде энергияға деген тапшылық
жағдай дамиды, мембраналардағы насостардың қызметтері одан
сайын
әлсірейді.
Мембраналардың
бүлінуі
нәтижесінде
митохондрийлар ыдырайды, ериді немесе ісінеді, олардың
тарақшалары
(кристалары)
бөлшектенеді,
жоғалады.
Митохондрийлардың ішінде бос қуыстар (вакуолдар) пайда
болады.
Жасуша ішіндегі ядроның мембраналары бұзылыстарынан
гендік ақпараттың сақталуы мен туынды жасушаларға берілуі,
нәруыз
түзілуі
бұзылады.
Ядрода
ДНҚ
молекуласының
репликациясы,
РНҚ-ның
транскрипциясы
өзгереді.
Жасушалардың әртүрлі бүліністерінде ядрода хроматиннің
тығыздығы, құрылымы және ядро ішінде тарауы, ядро түрінің
өзгеруі болады. Бұл кезде ядроның ісінуі, оның жиырылып,
бүрісіп қалуы (пикноз), жыртылуы және ыдырауы (кариорексис