Файл: Курсовая работа на тему Модель многосубстратного процесса микробного роста Простейшие кинетические схемы обобщенная модель многосубстратного микробного процесса.doc
Добавлен: 23.11.2023
Просмотров: 90
Скачиваний: 4
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
КУРСОВАЯ РАБОТА
на тему: Модель многосубстратного процесса микробного роста: Простейшие кинетические схемы; обобщенная модель многосубстратного микробного процесса
Введение
В микробиологии успех любого исследования во многом зависит от того, насколько изучены характер роста данного микроорганизма и его питательные потребности.
Начало выращиванию бактерий и выделению чистых культур положили работы отцов-основателей этой науки - Луи Пастера и Роберта Коха. К настоящему времени, спустя более чем 100 лет, получено в культуре и описано, вероятно, все еще менее одной десятой части существующих в природе видов прокариот.
О свойствах любой бактерии и ее роли в природе нельзя судить до тех пор, пока она не изучена в лабораторной культуре. Например, без этого часто невозможно определить, патогенен данный микроб или нет. Более чем 90% бактерий не выделены в виде культур, не описаны как виды и, таким образом, «скрыты» от человека; остается неизвестной их биологическая ценность. Выяснение питательных и физиологических потребностей необходимо не только для того, чтобы выращивать микроорганизмы. По требованиям к условиям культивирования и составу среды можно определить экологическую нишу и значение микроорганизма, а также его отличительные признаки, используемые для таксономии, экологического описания и решения практических задач. Характеристики роста отражают физиологические особенности микроорганизмов.
Наконец, управление ростом культур лежит в основе биотехнологического применения микроорганизмов. Все это подчеркивает важность изучения характера роста и питательных потребностей бактериальных культур.
1. Характер роста периодической культуры
Культуры называют периодическими, когда при росте в жидких средах в закрытых сосудах определенного объема микроорганизмы находятся в закрытой системе. Понятие микробный рост означает увеличение числа клеток микроорганизма. Под ростом отдельной бактериальной клетки понимают увеличение ее длины и последующее деление на две дочерние клетки (бинарное деление). У почкующихся бактерий возрастание числа клеток может также происходить в результате почкования. Дочерние клетки имеют те же размеры и состав, что и материнские. Когда время удвоения числа клеток остается постоянным, культура находится в экспоненциальной фазе роста (неверно называемой также логарифмической, или лог-фазой, по соображениям удобства построения графика роста в полулогарифмической шкале).
Удельная скорость роста определяется ферментативными возможностями данного вида бактерий, составом среды и всеми биофизическими факторами, влияющими на скорость роста. Как следствие, удельная скорость роста изменяется по экспоненциальному закону.
Исчерпание питательных веществ и накопление продуктов обмена до токсической концентрации приводят к постепенной остановке роста и переходу культуры в стационарную фазу, за которой следует фаза отмирания. Обычно причинами гибели - лизиса клеток в культуре становятся их энергетическое истощение и действие ферментов автолиза. Клетки некоторых бактерий не погибают после прекращения роста, благодаря тому что образуют споры или цисты, способны переживать неблагоприятные условия. В стационарной фазе происходит сложная адаптация клеток к условиям голодания и переживания и возможен скрытый рост выживших клеток на остатках лизированных клеток). Скорость роста в начальной стадии низка, поскольку в это время происходит адаптация клеток посевного материала к новым условиям.
Длительность этой фазы, называемой лаг-фазой или фазой задержки роста, зависит от количества клеток и возраста культуры, служившей посевным материалом, а также от различий в составе старой и новой сред. За это время клетки переходят из состояния адаптации к стационарной фазе роста культуры (часто именно такие, старые культуры используют для засева сред) в состояние активного роста. Длительность лаг-фазы увеличивается, если посевной материал был выращен на более богатой среде. При засеве материалом, взятым из культуры в экспоненциальной фазе роста, при одинаковом составе сред рост начинается без лаг-фазы.
В определенных условиях наблюдается не экспоненциальный, а линейный рост - если лимитирующий субстрат поступает в среду с постоянной скоростью, например добавляется в виде раствора по каплям или, в случае газа (такого, как О2), проникает в среду путем диффузии. Линейный рост может быть также обусловлен недостатком того или иного фермента, например при исчерпании его кофактора.
Микробные культуры используются во многих биотехнологических процессах в качестве биокатализаторов. Требуемый продукт они синтезируют чаще всего в стационарной фазе роста, поэтому вначале их выращивают отдельно в оптимальных для роста условиях (трофофаза) и затем переносят в «неростовые» условия, обеспечивающие синтез продукта, необходимого для данного технологического процесса (идиофаза).
Типичная кривая роста,
построенная в полулогарифмической шкале, приведена на рис. 1.
В периодической лабораторной культуре в течение короткого времени субстраты присутствуют в этом случае в среде в нелимитирующей концентрации; в природе же такие условия скорее редкое исключение, чем правило. Микробный рост в природе происходит в условиях, которые можно сравнить с концом фазы экспоненциального роста, когда количество субстратов становится лимитирующим и затем недостаточным для поддержания роста. Рост в таких условиях ограничен концентрацией субстрата, а не количеством клеточной массы.
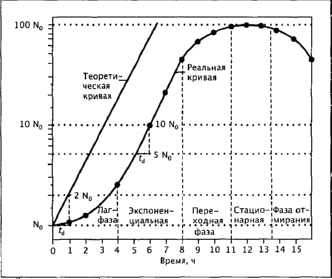
Рис. 1 - Типичная кривая роста периодической культуры, построенная в полулогарифмической шкале: зависимость логарифма числа клеток от времени (величина инокулята 1%). Для сравнения приведена теоретическая кривая экспоненциального роста
Кривая экспоненциального роста должна быть скорректирована введением понятия емкости с преобразованием в «логистическую» кривую роста.
где N - число клеток,
µ - скорость роста,
С - ёмкость, до которой возможен рост, определяемая, например, как количество лимитирующего рост субстрата.
Пока величина N невелика по сравнению с С, последний сомножитель близок к единице и несуществен. Однако, когда N приближается к соответствию С, экспоненциальный рост корректируется показателем емкости и нарастание биомассы прекращается с достижением стабильной конечной плотности популяции.
Показатель емкости С может относиться к лимитирующему субстрату, т.е. источнику углерода, азота, серы или фосфора, донору или акцептору электронов либо другим факторам. Согласно принципу Либиха, рост лимитирован субстратом, который исчерпывается первым, когда остальные могут присутствовать еще в избытке. Этот принцип не применим в полной мере к микробным клеткам, так как у них возможна определенная адаптация к изменениям условий (состава) среды. Содержание азота, серы или фосфора в клетках может варьировать в зависимости от количества субстрата в 2 - 10 раз.
Эффективность использования субстрата для достижения конечной емкости С определяется сродством к субстрату, Кв, данного организма. Этот термин означает концентрацию субстрата, при которой скорость роста равна половине максимальной. Если рост и потребление субстрата тесно сопряжены, К
в означает также концентрацию субстрата, при которой скорость потребления субстрата составляет 1/2 ее максимального значения. Значение Кв можно определить либо непосредственно по скорости поглощения субстрата (например, клеточной суспензией, образцом осадков или почвы), либо в лимитированной количеством субстрата хемостатной культуре при скорости разбавления, соответствующей 1/2 максимальной скорости роста.
2. Хемостат как модель роста микроорганизмов в природных системах
При хемостатном (непрерывном) культивировании, в отличие от периодического, в культиватор постоянно подается свежая питательная среда и концентрация одного из субстратов поддерживается на уровне, лимитирующем рост. Одновременно происходит постоянный отток культуры, так что объем ее в культиваторе не изменяется. Культиватор снабжен мешалкой для равномерного распределения свежей среды - рис. 2.
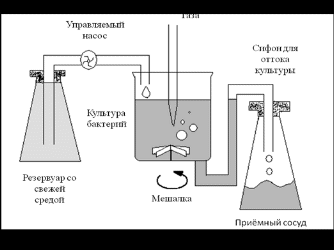
Рис. 2 - Система для непрерывного культивирования (хемостат)
Хемостат имитирует ситуацию, существующую в природе, в том отношении, что клетки поддерживаются в нем в условиях незначительного, но непрерывного поступления субстрата. Измеряемая концентрация субстрата не дает никакой информации о количестве субстрата, находящемся в обороте; она отражает лишь сродство к субстрату соответствующих микробных систем и его связь со скоростью поступления субстрата. Рост и оборот субстрата не обязательно связаны между собой стабильным соотношением, но оборот субстрата может быть все менее сопряжен с ростом по мере снижения скорости поступления субстрата.
Культивирование в хемостате:
обеспечивает постоянное получение молодых клеток, выросших в строго определенных условиях при известной удельной скорости роста; такие клетки необходимы для изучения различных регуляторных процессов;
позволяет определять важные показатели роста, а также изучать физиологическую реакцию культуры на лимитирование питательными веществами;
служит полезной моделью для описания роста во многих природных местообитаниях;
позволяет выявлять предпочтительно используемые субстраты в их смеси, а также конкуренцию или сосуществование двух или трех видов при данных условиях роста;
позволяет проводить подпитку токсичными субстратами в более высоких концентрациях;
позволяет исследовать стабильные двухкомпонентные культуры.
3. Способность микроорганизмов выживать в условиях голодания и при стрессе
В природе микроорганизмы обычно находятся в условиях недостатка субстрата или голодания, при которых важнее их способность к выживанию в таких условиях без существенного снижения плотности популяции, чем быстрый рост при избытке питательных веществ.
Бaктepии, растущие только при низких концентрациях субстратов, называют олиготрофными, отличая их от копиотрофных бактерий, способных расти только на богатых лабораторных средах. Олиготрофные бактерии трудно культивировать, поэтому об их физиологических и регуляторных способностях мало что известно; влияние на микроорганизмы условий голодания изучается главным образом на лабораторных штаммах таких бактерий, как Escherichia coli, Klebsiella или Paracoccus, т.е. не на тех организмах, для которых такие условия существования обычны в природных средах.
При изучении способов выживания микроорганизмов в природных условиях важно разделять различные типы голодания. Голодание по источнику углерода (обычно он служит одновременно и источником энергии) - это совершенно иная ситуация для клеток, чем голодание по источнику азота, серы или фосфора, препятствующее только росту, но не влияющее на процессы поддержания.
Клеточные стратегии выживания при недостатке энергетического субстрата сводятся кдвум основным типам, один из которых - это увеличение поглощения субстрата, даже при его низкой концентрации, и другой - снижение потребностей клеток в энергии. Оба эти варианта адаптации существуют в природных микробных популяциях, и возможно их сочетание.
Эффективность поглощения субстрата может возрастать благодаря увеличению числа систем транспорта (пермеаз и др.) либо индукции новых, более высокоаффинных транспортных систем, обеспечивающих поглощение субстрата при его низкой концентрации, например системы симпорта с использованием протондвижущей силы.
У многих хорошо изученных бактерий известны специфические высокоаффинные системы транспорта таких субстратов, как глицерол, фосфат, сахара и аминокислоты.
Другая стратегия адаптации - это увеличение поверхности клетки, например путем образования стебельков. Почкующиеся и стебельковые бактерий, например родов