ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 30.11.2023
Просмотров: 148
Скачиваний: 2
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
, то производится подкрепление. При ошибочном выборе пища не подается (наказание)
информационной триады. В рассматриваемом примере это соотношение нарушалось уменьшением времени за счет сокращения интервалов между отдельными сигналами. При этом не менялся уровень мотивации (который был высоким, так как животное было голодным) и не менялся объем информационной нагрузки - животным предъявлялось то же число условных раздражителей. Итак, животные осуществляли тот же объем аналитико-синтетической деятельности, но в условиях возникшего дефицита времени. Уже после нескольких предъявлений сигнальных раздражителей в этих условиях наблюдалось следующее изменение поведения: на включение условного сигнала животное сразу покидало стартовое место и подбегало к соответствующей кормушке, но возрастало время возвращения на стартовое место, и чем больше был дефицит времени, отведенного на решение всего объема задачи, тем медленнее возвращалось животное на стартовое место. Тем самым животное
само увеличивало интервал между условными сигналами, а значит, и время всего эксперимента, т.е. само устраняло дефицит времени и приводило в оптимальное для себя соотношение факторов информационной триады. Такое поведение значительно увеличивало период формирования патологических реакций, т.е. период предневроза, а в ряде случаев предотвращало развитие невроза.
Аналогичное поведение наблюдается, если меняется не время эксперимента между условными сигналами, а увеличивается нагрузка на аналитико-синтетическую деятельность мозга.
Понимание этих реакций как имеющих биологически положительное значение приобретает принципиальный характер: до недавнего времени они рассматривались как ранние симптомы патологии и соответственно подвергались подавлению («лечению») вместо того, чтобы принимать меры по их усилению. Описанные реакции отражают саморегуляционную деятельность мозга. Отсюда и большое практическое значение этих выводов - необходимо найти пути активации и усиления соответствующих саморегуляционных форм поведения. Это важно во всех периодах болезни, но особенно на стадии предболезни, когда саморегуляционные механизмы мозга хорошо выражены и их целенаправленное усиление может сыграть решающую роль в повышении психофизиологической резистентности организма.
Центральные механизмы защитной деятельности мозга.
После начала воздействия патогенного агента (например, неблагоприятного сочетания информационной триады) на высшую нервную деятельность в определенных структурах головного мозга развиваются патологические процессы, выявляемые электрофизиологическими, биохимическими и ультраструктурными исследованиями, которые нарушают нормальное взаимодействие между отдельными образованиями мозга (возникают новые корреляционные отношения между ними). После фиксации всех процессов в долгосрочной памяти формируются устойчивые патологические состояния мозга, что и проявляется внешне в различных неадаптивных реакциях.
Одновременно активируются структуры мозга, препятствующие развитию патологических процессов. Одним из внешних проявлений такой защитной активности мозга являются описанные выше саморегуляционные формы поведения. Кроме того, в мозгу развиваются еще два типа защитных реакций: происходит активация мозговых структур, неспецифически повышающих устойчивость нервной системы к внешним воздействиям, и активация мозговых структур, подавляющих формирование патологических процессов.
Эти выводы подтверждаются опытным путем с использованием методики локомоторной самостимуляции мозга. Для этого животным в разные образования головного мозга вживляются раздражающие электроды, через которые пропускается электрический ток. Включение тока производится самим животным во время его перемещения по полу, который разделен на участки. Каждый участок связан телеметрически с определенным раздражающим электродом, т.е. с определенной мозговой структурой. Как только животное становится на один из участков пола, включается стимуляция соответствующего образования мозга. Все описанное имеет место в помещении, где согласно ранее изложенному способу вырабатывается информационная патология высшей нервной деятельности (см. рис. 22-3).
Установлено, что если животное имеет возможность выбора между структурами для их самостимуляции, то на начальных стадиях развития информационной патологии высшей нервной деятельности оно преимущественно стимулирует прозрачную перегородку (рис. 22-4), т.е. вместо перемещения по полу экспе-
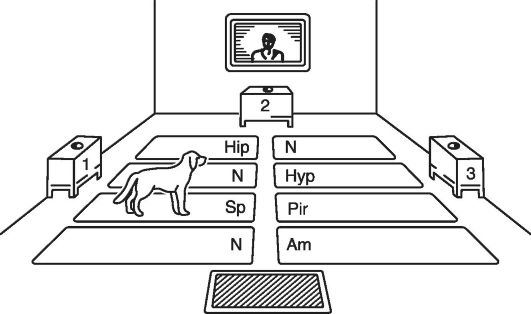 Рис. 22-4. Схема экспериментального помещения, приспособленного для выбора животным самостимуляции определенных структур мозга. N, Sp, Hyp, Hip, Pir, Am - участки поля, связанные с лимбическими структурами: N -
Рис. 22-4. Схема экспериментального помещения, приспособленного для выбора животным самостимуляции определенных структур мозга. N, Sp, Hyp, Hip, Pir, Am - участки поля, связанные с лимбическими структурами: N -
нейтральные для самостимуляции участки поля; Sp - septum pellucidum; Hyp - гипоталамус; Hip - гиппокамп; Pir - грушевидная извилина (lobus piriformis); Am - амигдолоидная область
риментальной комнаты в различных направлениях оно длительно (многие минуты) задерживается на участке, связанном с раздражением прозрачной перегородки. Такая активация этой структуры предотвращает развитие информационной патологии высшей нервной деятельности или существенно увеличивает период ее возникновения. Аналогичный эффект, хотя менее выраженный, отмечается при самостимуляции латерального гипоталамуса или медиального отдела миндалины. Очевидно, что прозрачная перегородка, латеральный гипоталамус и медиальная часть миндалины играют защитную функцию и препятствуют развитию патологии высшей нервной деятельности.
На ранних стадиях возникновения информационной патологии высшей нервной деятельности выявлены защитные (в том числе саморегуляционные) формы поведения и центральные механизмы, препятствующие ее развитию, т.е. обнаружено свойство головного мозга саморегулировать свое поведение в целях защиты организма от патогенного влияния факторов информационной триады. Установлены главные факторы риска возникновения информационной патологии высшей нервной деятельности и разработаны этиологически и патогенетически обоснованные принципы и методы ее профилактики и лечения.
22.7. ПОСТТРАВМАТИЧЕСКАЯ ПАТОЛОГИЯ
ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Этот вид патологии выражается в нарушении отдельных форм поведения. Наиболее подробно изучен патогенез нарушений пищевого, оборонительного, полового поведения, а также патогенез нарушения памяти, эмоций, цикла сон - бодрствование. Как отмечалось, создание моделей посттравматической патологии высшей нервной деятельности ставит своей целью воспроизведение различных повреждений мозга человека, вызванных кровоизлиянием в мозг, опухолью мозга, огнестрельным или другим травматическим ранением. Создание таких моделей достигается путем экстирпации мозговой ткани, перерезки проводящих путей, электрической коагуляцией отдельных участков мозга и др.
Нарушения отдельных форм поведения имеют некоторые общие
, а также частные проявления, типичные для каждой формы. Общими для всех являются нарушения баланса торможения и возбуждения.
Эти изменения могут завершиться глубоким угнетением функции неокортекса с последующим вовлечением в процесс подкорковых структур. В других случаях наблюдается усиление возбуждения. Изменения поведения общего характера во многом определяются типом или индивидуальными особенностями нервной системы.
22.7.1. Патология пищевого поведения
Она описана при обширных повреждениях неокортекса и частичных повреждениях лобной и орбитальной коры. При повреждениях (удалениях) лобных отделов неокортекса животные не способны выделить среди других съедобные предметы, поэтому поедают непригодные для еды вещи и не дифференцируют разные концентрации солевых растворов. Эти нарушения объясняются повреждением механизма афферентного формирования системы пищевого поведения. При повреждениях глазничной (орбитальной) области неокортекса резко снижается безусловно-рефлекторная секреция на пищевое раздражение, что объясняется наличием в этой области коркового представительства пищевого центра. При обширном удалении неокортекса или вследствие перерезки его проводящих путей происходят длительное снижение безусловно-рефлекторной пищевой секреции и почти полная потеря условных рефлексов, выработанных на основе пищевого подкрепления. Степень этих нарушений нарастает в восходящем эволюционном ряду животных и указывает на возрастающую роль неокортекса в регуляции пищевого поведения от низших животных к высшим.
Разнообразные отклонения от нормального пищевого поведения отмечаются при повреждении лимбических структур мозга. Так, у собак разрушение базально-латеральной части миндалевидного тела вызывает нарушение пищевых условных рефлексов и снижение безусловно-рефлекторной реакции на пищевые раздражители. Глубокие и характерные нарушения пищевого поведения возникают вследствие повреждения гипоталамуса; двустороннее разрушение вентромедиального ядра гипоталамуса у грызунов, хищных и приматов вызывает гиперфагию, а повреждение латерального гипоталамуса - афагию вплоть до гибели животных от кахексии.
Это дает основание считать, что патогенез описанных изменений пищевого поведения обусловлен нарушением регуляции чувства голода и насыщения. Между тем многочисленные наблюдения убеждают, что разные отделы гипоталамуса имеют отношение
к организации и регуляции и других компонентов пищевого поведения. Так, повреждение средней части латерального гипоталамуса вызывает нарушение исходного побуждения к пищедобыванию, а разрушение более латерального участка - нарушение регуляции приема пищи. Итак, нарушение разных звеньев пищевого поведения возникает при повреждении целого ряда структур головного мозга, объединенных в систему регуляции пищевого поведения, при этом неокортикальные структуры имеют первостепенное значение в формировании индивидуально приобретаемых пищевых реакций, а гипоталамические структуры играют исключительно важную роль в организации и регуляции безусловно-рефлекторного компонента разных звеньев пищевого поведения.
22.7.2. Патология оборонительного поведения
Вследствие экстирпации передних отделов лобной области новой коры у грызунов, хищных и приматов усиливается активнооборонительная реакция, которая иногда переходит в агрессивное поведение, ухудшает протекание условно-рефлекторных оборонительных реакций. Патогенез этих изменений связывается с ослаблением эмоции страха. Исчезновение реакции страха наблюдается и вследствие повреждения поясной извилины. Характерные изменения оборонительного поведения имеют место после повреждения миндалевидного тела - у животных исчезают страх и агрессивность, они становятся ручными.
Эти проявления травматического повреждения миндалевидного тела получили название синдрома Клювера-Бьюси. Вследствие повреждения вентромедиальной части гипоталамуса происходят усиление активнооборонительного поведения и возникновение агрессивности, тогда как повреждение задней части гипоталамуса усиливает пассивно-оборонительную реакцию - такие животные трусливы, их эмоциональные реакции протекают вяло. Предполагается, что патогенез описанных изменений оборонительной реакции имеет нейрохимическую природу и связан с нарушением регуляции серотонинергической системы мозга.
информационной триады. В рассматриваемом примере это соотношение нарушалось уменьшением времени за счет сокращения интервалов между отдельными сигналами. При этом не менялся уровень мотивации (который был высоким, так как животное было голодным) и не менялся объем информационной нагрузки - животным предъявлялось то же число условных раздражителей. Итак, животные осуществляли тот же объем аналитико-синтетической деятельности, но в условиях возникшего дефицита времени. Уже после нескольких предъявлений сигнальных раздражителей в этих условиях наблюдалось следующее изменение поведения: на включение условного сигнала животное сразу покидало стартовое место и подбегало к соответствующей кормушке, но возрастало время возвращения на стартовое место, и чем больше был дефицит времени, отведенного на решение всего объема задачи, тем медленнее возвращалось животное на стартовое место. Тем самым животное
само увеличивало интервал между условными сигналами, а значит, и время всего эксперимента, т.е. само устраняло дефицит времени и приводило в оптимальное для себя соотношение факторов информационной триады. Такое поведение значительно увеличивало период формирования патологических реакций, т.е. период предневроза, а в ряде случаев предотвращало развитие невроза.
| |
Аналогичное поведение наблюдается, если меняется не время эксперимента между условными сигналами, а увеличивается нагрузка на аналитико-синтетическую деятельность мозга.
Понимание этих реакций как имеющих биологически положительное значение приобретает принципиальный характер: до недавнего времени они рассматривались как ранние симптомы патологии и соответственно подвергались подавлению («лечению») вместо того, чтобы принимать меры по их усилению. Описанные реакции отражают саморегуляционную деятельность мозга. Отсюда и большое практическое значение этих выводов - необходимо найти пути активации и усиления соответствующих саморегуляционных форм поведения. Это важно во всех периодах болезни, но особенно на стадии предболезни, когда саморегуляционные механизмы мозга хорошо выражены и их целенаправленное усиление может сыграть решающую роль в повышении психофизиологической резистентности организма.
Центральные механизмы защитной деятельности мозга.
После начала воздействия патогенного агента (например, неблагоприятного сочетания информационной триады) на высшую нервную деятельность в определенных структурах головного мозга развиваются патологические процессы, выявляемые электрофизиологическими, биохимическими и ультраструктурными исследованиями, которые нарушают нормальное взаимодействие между отдельными образованиями мозга (возникают новые корреляционные отношения между ними). После фиксации всех процессов в долгосрочной памяти формируются устойчивые патологические состояния мозга, что и проявляется внешне в различных неадаптивных реакциях.
Одновременно активируются структуры мозга, препятствующие развитию патологических процессов. Одним из внешних проявлений такой защитной активности мозга являются описанные выше саморегуляционные формы поведения. Кроме того, в мозгу развиваются еще два типа защитных реакций: происходит активация мозговых структур, неспецифически повышающих устойчивость нервной системы к внешним воздействиям, и активация мозговых структур, подавляющих формирование патологических процессов.
| |
Эти выводы подтверждаются опытным путем с использованием методики локомоторной самостимуляции мозга. Для этого животным в разные образования головного мозга вживляются раздражающие электроды, через которые пропускается электрический ток. Включение тока производится самим животным во время его перемещения по полу, который разделен на участки. Каждый участок связан телеметрически с определенным раздражающим электродом, т.е. с определенной мозговой структурой. Как только животное становится на один из участков пола, включается стимуляция соответствующего образования мозга. Все описанное имеет место в помещении, где согласно ранее изложенному способу вырабатывается информационная патология высшей нервной деятельности (см. рис. 22-3).
Установлено, что если животное имеет возможность выбора между структурами для их самостимуляции, то на начальных стадиях развития информационной патологии высшей нервной деятельности оно преимущественно стимулирует прозрачную перегородку (рис. 22-4), т.е. вместо перемещения по полу экспе-
Рис. 22-4. Схема экспериментального помещения, приспособленного для выбора животным самостимуляции определенных структур мозга. N, Sp, Hyp, Hip, Pir, Am - участки поля, связанные с лимбическими структурами: N -
нейтральные для самостимуляции участки поля; Sp - septum pellucidum; Hyp - гипоталамус; Hip - гиппокамп; Pir - грушевидная извилина (lobus piriformis); Am - амигдолоидная область
риментальной комнаты в различных направлениях оно длительно (многие минуты) задерживается на участке, связанном с раздражением прозрачной перегородки. Такая активация этой структуры предотвращает развитие информационной патологии высшей нервной деятельности или существенно увеличивает период ее возникновения. Аналогичный эффект, хотя менее выраженный, отмечается при самостимуляции латерального гипоталамуса или медиального отдела миндалины. Очевидно, что прозрачная перегородка, латеральный гипоталамус и медиальная часть миндалины играют защитную функцию и препятствуют развитию патологии высшей нервной деятельности.
| |
На ранних стадиях возникновения информационной патологии высшей нервной деятельности выявлены защитные (в том числе саморегуляционные) формы поведения и центральные механизмы, препятствующие ее развитию, т.е. обнаружено свойство головного мозга саморегулировать свое поведение в целях защиты организма от патогенного влияния факторов информационной триады. Установлены главные факторы риска возникновения информационной патологии высшей нервной деятельности и разработаны этиологически и патогенетически обоснованные принципы и методы ее профилактики и лечения.
22.7. ПОСТТРАВМАТИЧЕСКАЯ ПАТОЛОГИЯ
ВЫСШЕЙ НЕРВНОЙ ДЕЯТЕЛЬНОСТИ
Этот вид патологии выражается в нарушении отдельных форм поведения. Наиболее подробно изучен патогенез нарушений пищевого, оборонительного, полового поведения, а также патогенез нарушения памяти, эмоций, цикла сон - бодрствование. Как отмечалось, создание моделей посттравматической патологии высшей нервной деятельности ставит своей целью воспроизведение различных повреждений мозга человека, вызванных кровоизлиянием в мозг, опухолью мозга, огнестрельным или другим травматическим ранением. Создание таких моделей достигается путем экстирпации мозговой ткани, перерезки проводящих путей, электрической коагуляцией отдельных участков мозга и др.
Нарушения отдельных форм поведения имеют некоторые общие
, а также частные проявления, типичные для каждой формы. Общими для всех являются нарушения баланса торможения и возбуждения.
Эти изменения могут завершиться глубоким угнетением функции неокортекса с последующим вовлечением в процесс подкорковых структур. В других случаях наблюдается усиление возбуждения. Изменения поведения общего характера во многом определяются типом или индивидуальными особенностями нервной системы.
| |
22.7.1. Патология пищевого поведения
Она описана при обширных повреждениях неокортекса и частичных повреждениях лобной и орбитальной коры. При повреждениях (удалениях) лобных отделов неокортекса животные не способны выделить среди других съедобные предметы, поэтому поедают непригодные для еды вещи и не дифференцируют разные концентрации солевых растворов. Эти нарушения объясняются повреждением механизма афферентного формирования системы пищевого поведения. При повреждениях глазничной (орбитальной) области неокортекса резко снижается безусловно-рефлекторная секреция на пищевое раздражение, что объясняется наличием в этой области коркового представительства пищевого центра. При обширном удалении неокортекса или вследствие перерезки его проводящих путей происходят длительное снижение безусловно-рефлекторной пищевой секреции и почти полная потеря условных рефлексов, выработанных на основе пищевого подкрепления. Степень этих нарушений нарастает в восходящем эволюционном ряду животных и указывает на возрастающую роль неокортекса в регуляции пищевого поведения от низших животных к высшим.
Разнообразные отклонения от нормального пищевого поведения отмечаются при повреждении лимбических структур мозга. Так, у собак разрушение базально-латеральной части миндалевидного тела вызывает нарушение пищевых условных рефлексов и снижение безусловно-рефлекторной реакции на пищевые раздражители. Глубокие и характерные нарушения пищевого поведения возникают вследствие повреждения гипоталамуса; двустороннее разрушение вентромедиального ядра гипоталамуса у грызунов, хищных и приматов вызывает гиперфагию, а повреждение латерального гипоталамуса - афагию вплоть до гибели животных от кахексии.
Это дает основание считать, что патогенез описанных изменений пищевого поведения обусловлен нарушением регуляции чувства голода и насыщения. Между тем многочисленные наблюдения убеждают, что разные отделы гипоталамуса имеют отношение
| |
к организации и регуляции и других компонентов пищевого поведения. Так, повреждение средней части латерального гипоталамуса вызывает нарушение исходного побуждения к пищедобыванию, а разрушение более латерального участка - нарушение регуляции приема пищи. Итак, нарушение разных звеньев пищевого поведения возникает при повреждении целого ряда структур головного мозга, объединенных в систему регуляции пищевого поведения, при этом неокортикальные структуры имеют первостепенное значение в формировании индивидуально приобретаемых пищевых реакций, а гипоталамические структуры играют исключительно важную роль в организации и регуляции безусловно-рефлекторного компонента разных звеньев пищевого поведения.
22.7.2. Патология оборонительного поведения
Вследствие экстирпации передних отделов лобной области новой коры у грызунов, хищных и приматов усиливается активнооборонительная реакция, которая иногда переходит в агрессивное поведение, ухудшает протекание условно-рефлекторных оборонительных реакций. Патогенез этих изменений связывается с ослаблением эмоции страха. Исчезновение реакции страха наблюдается и вследствие повреждения поясной извилины. Характерные изменения оборонительного поведения имеют место после повреждения миндалевидного тела - у животных исчезают страх и агрессивность, они становятся ручными.
Эти проявления травматического повреждения миндалевидного тела получили название синдрома Клювера-Бьюси. Вследствие повреждения вентромедиальной части гипоталамуса происходят усиление активнооборонительного поведения и возникновение агрессивности, тогда как повреждение задней части гипоталамуса усиливает пассивно-оборонительную реакцию - такие животные трусливы, их эмоциональные реакции протекают вяло. Предполагается, что патогенез описанных изменений оборонительной реакции имеет нейрохимическую природу и связан с нарушением регуляции серотонинергической системы мозга.
| |