Добавлен: 12.12.2023
Просмотров: 109
Скачиваний: 4
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
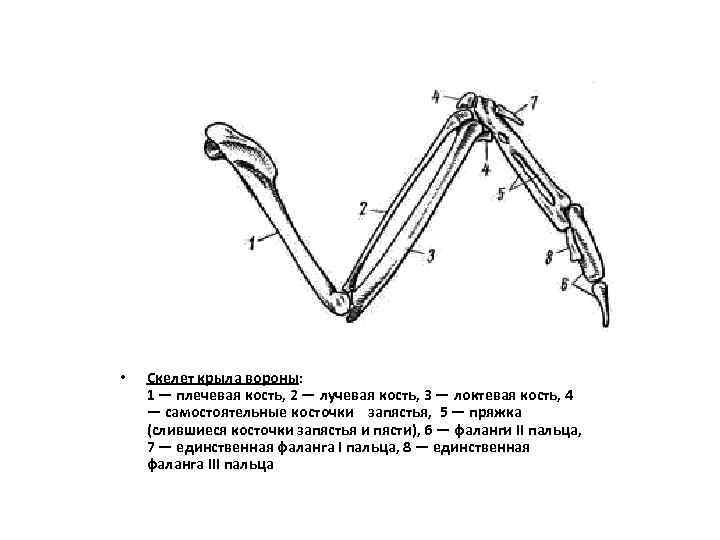
Особенно резкие изменения в связи с приспособлением к полету произошли в строении кисти. В проксимальном отделе запястья (carpus) сохраняются только две самостоятельные косточки (рис. 111, 4 ). Они связками почти неподвижно соединяются с костями предплечья. Подвижный сустав расположен между ними и остальными костями запястья, поэтому его называют интеркарпальным. Остальные кости запястья и все кости пясти (metacarpus) сливаются в единое образование— пряжку (carpometacarpus; рис. 7, 5 ). Резко редуцируется скелет пальцев. Сохраняются две фаланги II пальца (рис. 7, 6 ), продолжающие ось пряжки. От I пальца сохраняется одна фаланга (рис. 7, 7 ), к ней прикрепляется пучок перьев, образующий крылышко. III палец представлен также одной фалангой (рис. 7, 8 ), прикрепленной к основанию первой фаланги II пальца.
Эти преобразования в скелете кисти (образование carpometacarpus, редукция пальцев) обеспечивают прочную опору для первостепенных маховых перьев — части крыла, несущей в полете наибольшую нагрузку.
Тазовый пояс состоит из трех пар срастающихся вместе костей: Широкая и длинная подвздошная кость (ilium; см. рис. 6, 15 ; рис. 7, 4 ) на большом протяжении срастается со ложным крестцом. К ее наружному краю прирастает седалищная кость (ischium; см. рис. 6, 16 ; рис. 7, 5 ), а ниже расположена направленная назад палочкообразная лобковая кость (pubis; см. рис. 6, 17 ; рис. 7, 6 ). Все три кости таза образуют вертлужную впадину (acetabulum; см. рис. 6, 18 ), в которую входит головка бедра.
Большая поверхность тазовых костей и их прочное (неподвижное) соединение с осевым скелетом обеспечивают прочную опору для прикрепления мощных мышц. Эти особенности тазового пояса определяются тем, что передние конечности выполняют только функцию полета, а при движении по земле вся тяжесть тела поддерживается лишь задними конечностями.
Лобковые и седалищные кости правой и левой сторон не соединяются друг с другом на брюшной поверхности. Поэтому таз птиц называют открытым; он дает возможность откладывать крупные яйца. Можно предполагать, что открытый таз, увеличивающий подвижность брюшной стенки в тазовой области, способствует интенсификации дыхания в полете.
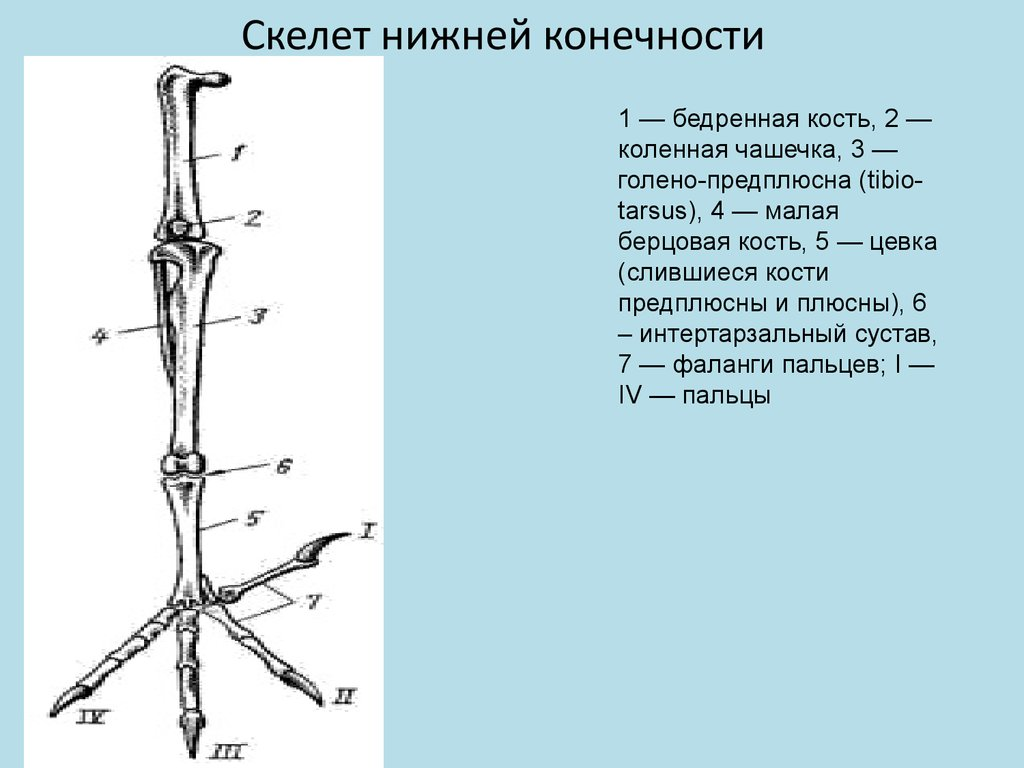
Скелет задней конечности представлен мощными трубчатыми костями. Бедро (femur; рис. 8, 1 ) на проксимальном конце имеет хорошо развитую округлую головку для сочленения с тазовым поясом. На дистальном конце образуются рельефные суставные поверхности для сочленения с костями голени. В области коленного сустава в мышечном сухожилии лежит коленная чашечка (patella; рис. 8, 2 ).
Основной элемент голени — массивная большая берцовая кость, с нижним дистальным концом которой сливаются две проксимальные кости предплюсны; образуется костный комплекс, представляющий собой голено-предплюсну, или тибио-тарзус (tibio-tarsus; рис. 8, 3 ). Малая берцовая кость (fibula; рис. 8, 4 ) сильно редуцирована и в виде маленькой тоненькой косточки прирастает к верхней части наружной поверхности большой берцовой кости.
Дистальные элементы предплюсны (tarsus) и все элементы плюсны (metatarsus) сливаются у птиц в единую кость — цевку, или плюсну-предплюсну (tarso-metatarsus; рис. 8, 5 ). Благодаря этому в конечности появляется добавочный рычаг. Подвижное сочленение располагается между двумя рядами костей предплюсны (между костями приросшими к голени, и костями, слившимися с плюсной), поэтому, как и у пресмыкающихся, этот сустав называется интертарзальным (рис. 8, 6 ).
На дистальном конце цевки образуются хорошо выраженные суставные поверхности для причленения фаланг пальцев (phalanges digitorum; рис. 8, 7 ). У подавляющего большинства птиц в задней конечности развиваются четыре пальца, из которых I направлен назад, а II, III, IV — вперед.
Мощные длинные кости скелета задних конечностей, появление добавочного рычага (образование tarso-metatarsus), резкий рельеф суставных поверхностей — все это обеспечивает прочность и подвижность задних конечностей.
 kkkk
kkkkЗаключение:
Морфологически класс птиц во многих чертах сохраняет большое сходство со своими предками — пресмыкающимися. На это указывает целый ряд признаков: почти полное отсутствие кожных желез, то, что дистальные отделы задних конечностей покрыты роговыми чешуями, роговой покров клюва, диапсидного типа череп (с редуцированной, однако, верхней дугой), имеющий только один затылочный мыщелок, состав костей черепа, то, что имеются шейные ребра и крючковидные отростки грудных ребер, открытый таз (как у некоторых ископаемых динозавров), большое сходство строения мочеполовой системы и периферических отделов кровеносной системы, присутствие клоаки, размножение откладыванием яиц, сходство эмбрионального развития и т. п.
В процессе эволюции птицы обособились от рептилиеобразных предков путем многих, но часто относительно небольших преобразований морфологических структур, обеспечиваюших существенную интенсификацию физиологических функций и тем самым резко повысивших общий уровень организации птиц по сравнению с пресмыкающимися. Полет и высокая общая подвижность птиц связаны с большим расходом энергии, требующим быстрой компенсации. Поэтому в эволюции птиц большое значение имело становление особенностей организации, связанных не только с обеспечением физической возможности полета, но и с общим повышением интенсивности обмена веществ.
Возможность полета птиц обеспечивается целым рядом прямых приспособлений. К ним относятся: компактность тела, жесткость и неподвижность туловищного отдела осевого скелета и редукция его хвостового отдела, резкое увеличение размеров грудины и образование на ней киля, прочность пояса передних конечностей (в том числе и образование специального амортизирующего устройства — вилочки), строение скелета крыла (включая и особенности суставных поверхностей и особенно резкие изменения в строении кисти), мощность мускулатуры, обеспечивающая движения крыла, развитие перьев — легких и прочных роговых образований, создающих несущие плоскости крыльев и хвоста и обеспечивающих обтекаемость тела в по лете, редукция зубов, облегчающая скелет черепа, общее увеличение объема головного мозга и особенно сильное развитие мозжечка — центра равновесия и координации движений, увеличение массы нервных клеток спинного мозга в области крыльев и т. д.
олее высокий уровень организации птиц, по сравнению с пресмыкающимися, находит свое отчетливое выражение не только в значительно более широком их распространении по всему земному шару, но и в более сложных и разнообразных взаимоотношениях со средой, с условиями жизни. Так, при сезонных неблагоприятных изменениях условий жизни (наступление зимы) пресмыкающиеся и земноводные прекращают свою активность, впадают в состояние оцепенения. У птиц в целом реакции при наступлении неблагоприятных сезонов года многообразнее и шире. Часть птиц совершает миграции (протяженностью до 5—10 тыс. км и более), улетая на зимовки в благоприятные по погодным и кормовым условиям районы. Однако значительное число видов остается в умеренных и северных широтах в течение всего года. Переживать суровые условия зимы им помогают довольно разнообразные приспособления.
Для многих видов свойственна резко выраженная сезонная смена кормов: переход зимой на питание менее калорийными, но массовыми кормами (например, тетерев летом питается насекомыми, ягодами, зелеными частями растений, зимой — преимущественно почками и сережками некоторых пород деревьев). Неперелетные (оседлые) птицы совершают небольшие перемещения, концентрируясь в участках с большим количеством пищи (глухарь в сосновых лесах, где питается хвоей сосны; синицы, вороны — в населенных пунктах и т. п.). Некоторые виды осенью запасают пищу, пряча ее в укромных местах (кедровки, сойки, поползни — орехи лещины, «орешки» кедра, желуди; синицы — семена деревьев и кустарников). Оседлые птицы во время осенней линьки получают перьевой покров с лучшими теплоизолирующими свойствами, чем летний (большее развитие пуховой части пера, большее число перьев и пуха на единицу поверхности тела и т. п.); у некоторых видов
зимний наряд резко отличается по окраске, помогая маскировке (белая куропатка).
У части видов отчетливо выражены и приспособительные особенности поведения: ночевки в укрытиях (тетеревинные и некоторые другие птицы — в снегу; мелкие воробьиные — в дуплах, щелях за корой и т. п.). При этом птицы некоторых видов в сильные морозы ночуют группами, взаимно обогревая в убежище друг друга (синицы, воробьи и др.).
Несмотря на резкое повышение уровня жизнедеятельности и широкие экологические возможности, открываемые гомойотермией, отчетливая специализация птиц к полету в известной мере ограничивает круг экологических приспособлений этих животных. Отсутствие столь узкой специализации у млекопитающих создало предпосылки к возникновению в процессе эволюции этого класса значительно большего разнообразия жизненных форм и более многообразных взаимоотношений со средой.