ВУЗ: Московский государственный университет технологий и управления им. К.Г. Разумовского
Категория: Книга
Дисциплина: Медицина
Добавлен: 09.02.2019
Просмотров: 24738
Скачиваний: 28

Рис. 5.
типичного синапса.
Объяснение в тексте.
Более подробно эти процессы
рассматриваются в разделе "Ней-
рохимические системы мозга".
Рецепторы нейронов — это
белковые структуры, располо-
женные на внешней поверхности
мембраны клеток. Они способны
"распознавать" и связывать биоло-
гически активные вещества —
различные эн-
догенные вещества, а также экзо-
генные соединения, в том числе
психофармакологические средст-
ва. Соединения, которые могут свя-
зывать рецепторы, называются ли-
гандами. Лиганды бывают эндо-
генными и экзогенными.
Распознавание лиганда ре-
цептором обеспечивается специ-
альными структурными элемен-
тами, или сайтами. Специфич-
ность связывания лиганда происходит благодаря структурному соответствию
молекул лиганда и рецептора, когда они подходят друг к другу по типу "ключ
к замку". Реакция связывания является моментом запуска каскада внутри-
клеточных реакций, приводящих к изменению функционального состояния
нейрона. В зависимости от "силы" и "прочности" связывания лиганда с
рецептором употребляют понятие аффинности (сродства) лиганда по отно-
шению к рецептору.
При связывании рецептора с
может происходить как актива-
ция, так и блокада рецептора. В связи с этим говорят об агонистах и
антагонистах рецепторов, а также о частичных агонистах (рис. 7).
Максимальную эффективность в отношении активации рецептора
имеет полный агонист, минимальную (практически нулевую) — антагонист.
Между ними находятся вещества, называемые частичными агонистами. Пос-
ледние действуют значительно мягче, чем полные агонисты. Частичные
агонисты, кроме того, занимая определенное пространственное положение
в молекуле рецептора, могут предотвращать избыточное действие полного
агониста, т.е. действуют частично как антагонисты. В этом случае употреб-
ляют понятие
Высокой аффинностью могут обладать как агонисты, так и антагонисты
рецептора. Агонист активирует рецептор, вызывая соответствующий физио-
логический эффект, в то время как антагонист, связываясь с рецептором,
блокирует его и предотвращает развитие физиологического эффекта, вы-
являемого агонистами. Примером антагонистов могут служить нейролеп-
тики, которые предотвращают эффекты дофамина на уровне дофамино-
вого рецептора.
При связывании лиганда с рецептором происходит изменение конфи-
гурации последнего (рис. 7).
Многие вещества, как эндогенные, так и экзогенные, реагируют не с
одним, а с несколькими типами рецепторов — "семейством" их, которое
подразделяется на отдельные типы. Примером могут служить многие ней-
114
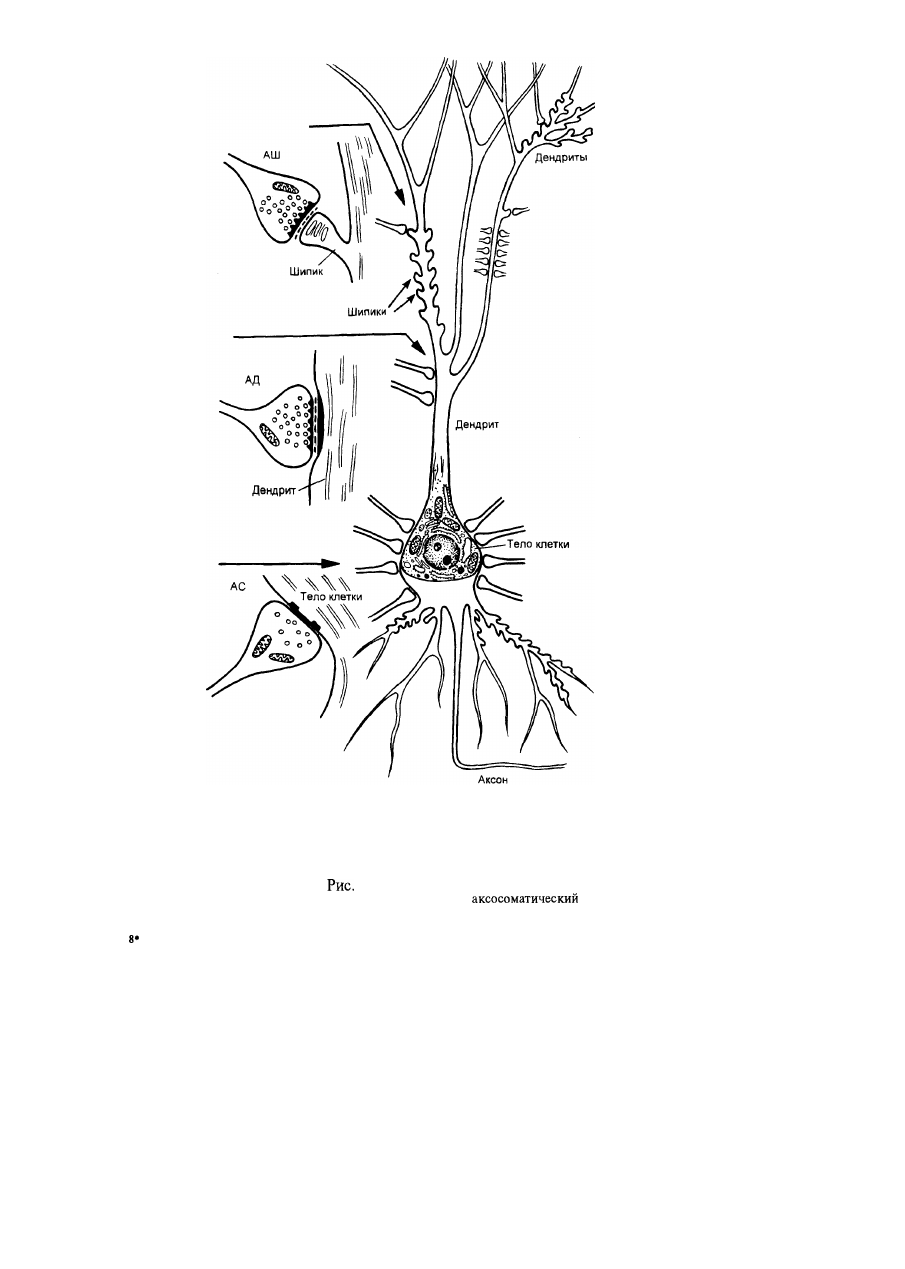
6. Типы синапсов.
Аксошипиковый (АШ); аксодендритный (АД);
(АС).
115

ротрансмиттеры, реагирующие с не-
сколькими типами специфических ре-
цепторов (например,
до-
фаминовых рецепторов). Существова-
ние нескольких рецепторов к одному
лиганду носит название гетерогенности
рецепторов.
Представление о функции рецеп-
торов было бы неполным, если не
представить внутриклеточные процес-
сы, развивающиеся после связывания
рецептора соответствующим вещест-
вом, и механизмы, обеспечивающие
трансформацию внешнего сигнала в
процессы, приводящие к появлению
нервного импульса. Связывание ли-
ганда с рецептором может приводить
либо непосредственно к открытию (или
закрытию) соответствующих ионных
каналов (см. рис. 7), либо к активации
вторичных мессенджерных систем (в ка-
честве первичного мессенджера рас-
сматривается вещество, реагирующее с
рецептором).
Первые упоминания о вторичных
мессенджерных системах появились в
связи с работами E.Sutherland и соавт.
(1950), которые показали, что адрена-
лин стимулирует гликогенез путем уве-
личения концентрации циклического
(цАМФ) в клет-
ке. Оказалось, что этот вторичный мес-
сенджер опосредует и другие клеточ-
ные реакции. В дальнейшем была вы-
явлена связь действия цАМФ с акти-
вацией белковых киназ — ферментов, фосфорилирующих белки, что при-
водит к изменению их структуры и активности.
Позднее были открыты и другие вторичные мессенджеры. Сейчас вы-
деляют среди них 3 класса: 1) циклические нуклеотиды (цАМФ, цикличес-
кий гуанозинмонофосфат — цГМФ); 2) ионы кальция
3) метаболи-
ты фосфолипидов — инозитол-1,4,5-трифосфат
диглицерин (ДАГ),
арахидоновую кислоту. В отличие от других вторичных мессенджеров
транспортируется в нейрон из внутриклеточного пространства.
Мембраны нейрона содержат специализированные трансмембранные
белки, которые формируют ионные каналы не только для
но и для
других ионов, концентрация которых по обе стороны мембраны влияет на
изменение мембранного потенциала. Происходят поляризация и деполяри-
зация мембраны, т.е. изменение трансмембранного потенциала. Наибольшее
значение в этих процессах имеют ионные каналы для натрия
калия
хлора
и кальция
Рис. 7. Взаимодействие рецептора с
агонистом, частичным агонистом и
антагонистом на примере холинерги-
ческого рецептора с тетраметрической
структурой.
116

НЕЙРОХИМИЧЕСКИЕ СИСТЕМЫ МОЗГА
В
годах с помощью гистохимических и радиоиммунологических
методов было установлено, что в мозге имеются дифференцированные ней-
рохимические системы — норадренергическая, дофаминергическая, серото-
нинергическая и др. Они представляют собой комплекс мозговых структур,
функция которых определяется наличием общего нейротрансмиттера и ре-
цепторов, взаимодействующих с ним. В одних из структур, входящих в
нейрохимическую систему, расположены тела нейронов, в других оканчи-
ваются терминали нервных клеток. В последнем случае говорят о
на те или иные мозговые образования. В одних и тех же структурах
мозга могут располагаться клетки и проекции нескольких нейрохимических
систем.
Дофаминергическая система. В этой системе мозга различают 7 отдель-
ных подсистем (систем, трактов): нигростратную, мезокортикальную, мезо-
лимбическую, тубероинфундибулярную,
диенце-
фалоспинальную и ретинальную. Из них первые 3 являются основными
(рис. 8). Тела нейронов нигростриатной, мезокортикальной и мезолимби-
ческой систем расположены на уровне среднего мозга, образуют комлекс
нейронов черной субстанции и вентрального поля покрышки. Они состав-
ляют непрерывную клеточную сеть, проекции которой частично перекры-
ваются, поскольку аксоны этих нейронов идут вначале в составе одного
крупного тракта (медиального пучка переднего мозга), а оттуда расходятся
в разные мозговые структуры. Формирование нигростриатной, мезолимби-
ческой и мезокортикальной систем определяется областями, где оканчива-
ются аксоны дофаминергических нейронов, т.е. локализацией их проекций.
Некоторые авторы объединяют мезокортикальную и мезолимбическую под-
системы в единую систему. Более обоснованным является выделение мезо-
кортикальной и мезолимбической подсистем соответственно проекциям в
лобную кору и лимбические структуры мозга (см. рис. 8).
система. Нигростриатный тракт является самым мощ-
ным в дофаминергической системе мозга. Аксонами нейронов этого тракта
выделяется около 80 % мозгового дофамина. Тела дофаминовых нейронов,
образующих этот путь, находятся в основном в компактной части черной
субстанции, но часть волокон берет начало также от нейронов латерального
отдела вентрального поля покрышки среднего мозга.
Клетки компактной части черной субстанции дают проекции в дорсаль-
ный стриатум (полосатое тело), а клетки вентрального поля покрышки — в
вентральный стриатум. Наиболее плотно расположены дофаминергические
волокна в стриатуме — они начинаются от латеральных отделов черной
субстанции того же полушария. Эти волокна оканчиваются на нейронах
хвостатого ядра и скорлупы, т.е. в неостриатуме. Дофаминергическую ин-
нервацию получают также другие структуры, в частности базальные ган-
глии — бледный шар (палеостриатум) и субталамическое ядро. У хвостатого
ядра более плотная иннервация отмечается в головке и значительно меньше
плотность дофаминергических проекций в каудальной части.
система. Тела нейронов, образующих мезокортикаль-
ный тракт, находятся в вентральной части покрышки среднего мозга, а
основные проекции этих нейронов достигают лобной (преимущественно
префронтальной, поле 10 по Бродману — рис. 9) коры. Соответствующие
окончания расположены в основном в глубоких слоях лобной коры
Мезокортикальная дофаминовая система оказывает большое влияние на
117

активность нейронов, образующих корково-корковые, корково-таламичес-
кие и корково-стриатные пути.
система. Источники дофаминергических проекций,
т.е. тела нейронов этой системы, расположены в вентральном поле покрыш-
ки среднего мозга и частично в компактной части черной субстанции. Их
отростки идут в поясную извилину, энториальную кору, миндалину, обоня-
тельный бугорок, аккумбентное ядро, гиппокамп, парагиппокампальную
извилину, перегородку и другие структуры лимбической системы мозга.
Имея обширные связи, мезолимбическая система опосредованно проециру-
ется также на лобную кору и гипоталамус. Это определяет широкие функции
системы, которая участвует в механизмах памяти, эмоций,
обучения и
регуляции.
Другие тракты. Тубероинфундибулярный тракт образован аксонами
нейронов, расположенных в аркуатном ядре гипоталамуса. Отростки таких
нейронов достигают наружного слоя срединного возвышения. Этот тракт
осуществляет контроль секреции пролактина. Дофамин тормозит его секре-
цию и поэтому содержание пролактина в плазме крови служит косвенным
показателем функции дофаминергической системы мозга, что часто исполь-
зуют для оценки влияния на нее психофармакологических средств. Инцер-
тракт начинается от zona incerta и оканчивается в
дорсальном и переднем отделах медиального таламуса, а также в перивент-
рикулярной области. Он принимает участие в нейроэндокринной регуляции.
Источником проекций диэнцефалоспинального тракта являются нейроны
заднего гипоталамуса, отростки которых достигают задних рогов спинного
мозга. Ретинальный тракт расположен в пределах сетчатки глаза. Особен-
ности этого тракта делают его среди других дофаминергических трактов
достаточно автономным.
Приведенная дифференциация дофаминергических образований мозга
не является абсолютной, так как проекции дофаминергических нейронов
разных трактов "перекрываются". Кроме того, в мозге отмечается и диф-
фузное распределение дофаминергических элементов (отдельных клеток с
отростками).
Дофаминергические системы мозга созревают преимущественно в по-
станальном периоде.
Дофаминовые терминали образуют синапсы преимущественно на ши-
пиках и стволах дендритов — это аксошипиковые и аксодендритные синап-
сы (их более 90 %). Лишь единичные синапсы (менее 10 %) расположены
на телах нейронов (аксосоматические) и на аксонах (аксоаксональные).
Основными типами дофаминовых рецепторов являются Д1- и
цепторы. Недавно были открыты также рецепторы ДЗ, Д4 и Д5. Они все
находятся главным образом на постсинаптической мембране. Но в дофами-
нергической системе существуют также ауторецепторы, расположенные на
теле нейронов, аксонах, дендритах и терминалях, которые реагируют на
собственный дофамин, регулируя его синтез и выделение. Их стимуляция
приводит к снижению активности дофаминовых нейронов.
Большая часть охарактеризованных дофаминовых рецепторов относится
к
О функции
известно меньше. Их от Д2-
рецепторов отличает способность стимулировать активность фермента аде-
нилатциклазы, которая в свою очередь участвует в синтезе второго мессенд-
цАМФ. Д1- и Д2-рецепторы существуют в двух
высоко-
и низкоаффинной, что определяется по их способности к связыванию
агонистов и антагонистов. Было показано, что Д2-рецепторы имеют высокое
118