Файл: Курс лекций, нгу 2000. Лекция Что такое молекулярная биология. Доказательства роли нуклеиновых кислот.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 11.01.2024
Просмотров: 623
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
В  ведение в молекулярную биологию Дымшиц Г.М. и др.Курс лекций, НГУ 2000.
ведение в молекулярную биологию Дымшиц Г.М. и др.Курс лекций, НГУ 2000.
Определение предмета молекулярная биологияТермин "молекулярная биология" принадлежит Фрэнсису Крику, которому надоело в ответ на вопрос о его профессии объявлять себя смесью кристаллографа, биохимика, биофизика и генетика. После атомной бомбежки Хиросимы и Нагасаки в 1945г. началось бегство ученых из физики, а в 1947г. Нобелевский лауреат физик Эрвин Шредингер написал книгу "Что такое жизнь с точки зрения физика?", которая привлекла в биологию многих физиков и математиков. Определение: Mолекулярная биология - это наука о механизмах хранения, воспроизведения, передачи и реализации генетической информации, о структуре и функциях нерегулярных биополимеров - нуклеиновых кислот и белков. Начав с изучения биологических процессов на молекулярно-атомном уровне, молекулярная биология перешла к сложным надмолекулярным клеточным структурам, а в настоящее время успешно решает проблемы генетики, физиологии, эволюции и экологии.Основные этапы развития молекулярной биологии1. Романтический период 1935-1944гг. Макс Дельбрюк и Сальвадор Лурия занимались изучением репродукции фагов и вирусов, представляющих собой комплексы нуклеиновых кислот с белкамиВ 1940г. Джордж Бидл и Эдуард Татум сформулировали гипотезу - "Один ген - один фермент". Однако, что такое ген в физико-химическом плане тогда еще не знали. 2. Второй романтический период1944-1953гг. Была доказана генетическая роль ДНК. В 1953 г. появилась модель двойной спирали ДНК, за которую ее создатели Джеймс Уотсон, Френсис Крик и Морис Уилкинс были удостоены Нобелевской премии. 3. Догматический период1953-1962гг. Сформулирована центральная догма молекулярной биологии:Перенос генетической информации идет в направлении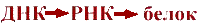 В 1962 г. был расшифрован генетический код. 4. Академический период с 1962г. по настоящее время, в котором с 1974 года выделяют генно-инженерный подпериод.Основные открытия
В 1962 г. был расшифрован генетический код. 4. Академический период с 1962г. по настоящее время, в котором с 1974 года выделяют генно-инженерный подпериод.Основные открытия
Доказательства генетической роли нуклеиновых кислот
Бескапсульные невирулентные пневмококки приобрели некоторые свойства убитых капсульных бактерий.Произошла трансформация.Определение: Трансформация - это приобретение одним организмом некоторых признаков другого организма за счет захвата части его генетической информации. В 1944г. этот эксперимент был повторен Освальдом Эйвери, Колином Мак-Леодом и Маклином Мак-Карти в варианте смешивания бескапсульных пневмококков с взятыми от капсульных белками, полисахаридами или ДНК. В результате этого эксперимента была выявлена природа трансформирующего фактора.
Трансформирующим фактором оказалась ДНК. 2. 1952г. Эксперимент Альфреда Херши и Марты Чейз.Фаги (бактериофаги) - это вирусы, размножающиеся в бактериях.E. сoli - кишечная палочка (эубактерия). Суть опыта: фаги, у которых белковая оболочка была мечена радиоактивной серой (S35), а ДНК - радиоактивным фосфором (Р32), инкубировали с бактериями. Затем бактерии отмывали. В смывных водах не обнаруживали Р32, а в бактериях - S35 Следовательно, внутрь попала только ДНК. Через несколько минут из бактерии выходили десятки полноценных фагов, содержащих и белковую оболочку, и ДНК.
Отсюда следовал однозначный вывод о том, что именно ДНК выполняет генетическую функцию - несет информацию как о создании новых копий ДНК, так и о синтезе фаговых белков. 3. 1957г. Опыты Френкеля - Конрата Френкель-Конрат работал с вирусом табачной мозаики (ВТМ). В этом вирусе содержится РНК, а не ДНК. Было известно, что разные штаммы вируса вызывают разную картину поражения листьев табака. После смены белковой оболочки "переодетые" вирусы вызывали картину поражения, характерную для того штамма, чья РНК была покрыта чужим белком.
Следовательно, не только ДНК, но и РНК может служить носителем генетической информации.На сегодняшний день существуют сотни тысяч доказательств генетической роли нуклеиновых кислот. Приведенные три являются классическими.
-
 Лекция 1. Что такое молекулярная биология. Доказательства ролинуклеиновых кислот.
Лекция 1. Что такое молекулярная биология. Доказательства ролинуклеиновых кислот. -
Определение предмета молекулярная биология -
Основные этапы развития-
Первый романтический период -
Второй романтический период -
Догматический период -
Академический период
-
-
Основные открытия -
Доказательства генетической роли нуклеиновых кислот-
Опыты Ф. Гриффита -
Эксперимент А.Херши и М.Чейз -
Опыты Френкеля - Конрата
-
-
Вопросы к лекции 1
-
- Лекция 2. Строение нуклеиновых кислот
-
Хронология открытий, подготовивших создание Уотсоном и Криком модели двойной спирали ДНК -
Строение нуклеиновых кислот. Мономеры. -
Принципы строения ДНК -
Формы двойной спирали ДНК -
Особенности строения и виды РНК-
Отличия между ДНК и РНК -
Виды РНК
-
-
Функции ДНК -
Вопросы к лекции 2
-
- Лекция 3. Структура белков
-
Аминокислоты-
Структура аминокислот -
Классификация аминокислот
-
-
Первичная структура белка -
Вторичная структура белка -
Третичная структура белка -
Четвертичная структура белка. -
Серповидно-клеточная анемия, как пример влияния первичной структуры на третичную и четвертичную -
Глобуллярные и фибриллярные белки -
Вопросы к лекции 3
-
- Лекция 4. Функции белков
-
Вопросы к лекции 4
-
- Лекция 5. Генетический код
-
Свойства генетического кода -
Свойства генетического кода (продолжение) -
Вопросы к лекции 5
-
- Лекция 6. Транскрипция
-
Принципы транскрипции -
Понятие об опероне -
Субъединичный состав РНК-полимеразы Е. сoli -
Особенности структуры промотора -
Этапы транскрипции-
Узнавание и прочное связывание -
Инициация -
Элонгация -
Терминация
-
-
Ингибиторы транскрипции прокариот -
Вопросы к лекции 6
-
- Лекция 7. Транскрипция и трансляция у прокариот
-
Регуляция транскрипции у прокариот-
Схема негативной индукции Жакоба и Моно -
Схема позитивной индукции -
Схема позитивной репрессии -
Схема негативной репрессии -
Позитивный контроль работы lac-оперона
-
-
Вопросы к лекции 7
-
- Лекция 8. Синтез белка в клетке. Подготовительный этап
-
Синтез белка в клетке-
Структура транспортной РНК -
Рекогниция -
Структура рибосом
-
-
Вопросы к лекции 8
-
- Лекция 9. Синтез белка на рибосоме
-
Механизм трансляции на прокариотических рибосомах -
Регуляция образования рибосомных РНК и белков рибосом E.сoli -
Аттенуация -
Вопросы к лекции 9
-
- Лекция 10. Транскрипция и трансляция у эукариот
-
Эукариотические РНК - полимеразы -
Образование рибосом у эукариот -
Особенности транскрипции у эукариот -
Вопросы к лекции 10
-
- Лекция 11. Процессинг mРНК
-
Этапы процессинга mРНК -
Кепирование -
Полиаденилирование -
Сплайсинг-
Альтернативный сплайсинг -
Автосплайсинг
-
-
Малые РНК -
Редактирование -
Вопросы к лекции 11
-
- Лекция 12. Репликация. Принципы. Синтез ДНК in vitro
-
Принципы репликации -
Доказательство полуконсервативного характера репликации -
Ферментативная система синтеза ДНК in vitro -
Понятие о матрице и затравке -
Вопросы к лекции 12
-
- Лекция 13.Схемы репликации ДНК in vivo
-
Схемы репликации-
Схема непрерывной антипараллельной репликации in vivo по Корнбергу -
Схема непрерывной параллельной репликации Джона Кэрнса
-
-
Сравнительные характеристики ДНК-полимераз E. сoli -
Схема прерывистой антипараллельной репликации Рейджи Оказаки -
Инициация репликации -
Вопросы к лекции 13
-
- Лекция 14. Топология репликации
-
Проблемы денатурации ДНК матрицы -
Геликазы -
Топоизомеразы -
Современная схема репликации ДНК E. сoli -
Вопросы к лекции 14
-
- Лекция 15. Особенности репликации ДНК эукариот
-
Полирепликонность -
Проблема репликации концов линейных молекул -
Вопросы к лекции 15
-
- Лекция 16. Репарация
-
Причины ошибок при синтезе ДНК -
Этапы проверки точности синтеза ДНК -
Основные репарабельные повреждения в ДНК и принципы их устранения-
Апуринизация -
Дезаминирование -
Тиминовые димеры
-
-
Вопросы к лекции 16
-
- Лекция 17. Структура генома
-
Размер генома -
"Избыточность" эукариотического генома -
Компактность эукариотического генома-
Общая характеристика гистонов -
Четыре уровня компактизации ДНК
-
-
Вопросы к лекции 17
-
- Лекция 18. Классификация генов в геноме
-
Основы метода ренатурации ДНК -
Быстрые повторы -
Умеренные повторы -
Уникальные гены -
Классификация генов -
Вопросы к лекции 18
-
- Лекция 19. Нестабильность генома. Обратная транскрипция.
-
Классы мобильных генетических элементов-
IS- элементы -
Tn-транспозоны -
Умеренные фаги -
Эффекты, вызываемые мобильными элементами -
Молекулярные основы канцерогенеза
-
-
Вопросы к лекции 19
-
- Лекция 20. Молекулярно-биологические основы возникновения жизни на Земле
-
Гипотезы возникновения жизни -
Теория биопоэза -
Эволюция пробионтов -
Вопросы к лекции 20
-
-
Список терминов -
Список рисунков -
Поиск по сайту -
Заключение -
Литература
Определение предмета молекулярная биологияТермин "молекулярная биология" принадлежит Фрэнсису Крику, которому надоело в ответ на вопрос о его профессии объявлять себя смесью кристаллографа, биохимика, биофизика и генетика. После атомной бомбежки Хиросимы и Нагасаки в 1945г. началось бегство ученых из физики, а в 1947г. Нобелевский лауреат физик Эрвин Шредингер написал книгу "Что такое жизнь с точки зрения физика?", которая привлекла в биологию многих физиков и математиков. Определение: Mолекулярная биология - это наука о механизмах хранения, воспроизведения, передачи и реализации генетической информации, о структуре и функциях нерегулярных биополимеров - нуклеиновых кислот и белков. Начав с изучения биологических процессов на молекулярно-атомном уровне, молекулярная биология перешла к сложным надмолекулярным клеточным структурам, а в настоящее время успешно решает проблемы генетики, физиологии, эволюции и экологии.Основные этапы развития молекулярной биологии1. Романтический период 1935-1944гг. Макс Дельбрюк и Сальвадор Лурия занимались изучением репродукции фагов и вирусов, представляющих собой комплексы нуклеиновых кислот с белкамиВ 1940г. Джордж Бидл и Эдуард Татум сформулировали гипотезу - "Один ген - один фермент". Однако, что такое ген в физико-химическом плане тогда еще не знали. 2. Второй романтический период1944-1953гг. Была доказана генетическая роль ДНК. В 1953 г. появилась модель двойной спирали ДНК, за которую ее создатели Джеймс Уотсон, Френсис Крик и Морис Уилкинс были удостоены Нобелевской премии. 3. Догматический период1953-1962гг. Сформулирована центральная догма молекулярной биологии:Перенос генетической информации идет в направлении
-
1944г.
Доказательство генетической роли ДНК. Освальд Эйвери, Колин Мак-Леод, Маклин Мак-Карти
1953г.
Установление структуры ДНК. Джеймс Уотсон, Френсис Крик
1961г.
Открытие генетической регуляции синтеза ферментов. Андре Львов, Франсуа Жакоб, Жак Моно
1962г.
Расшифровка генетического кода. Маршалл Нирнберг, Генрих Маттеи, Северо Очоа
1967г.
Синтез in vitro биологически активной ДНК. Артур Корнберг (неформальный лидер молекулярной биологии)
1970г.
Химический синтез гена. Гобинд Корана
1970г.
Открытие фермента обратной транскриптазы и явления обратной транскрипции. Говард Темин, Дэвид Балтимор, Ренато Дульбеко
1974г.
Открытие рестриктаз. Гамильтон Смит, Даниэль Натанс, Вернер Арбер
1978г.
Открытие сплайсинга. Филипп Шарп
1982г.
Открытие автосплайсинга. Томас Чек
Доказательства генетической роли нуклеиновых кислот
-
1928г. Опыты Фредерика Гриффита.
| Опыты Гриффита | |
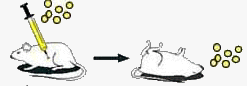 | После заражения пневмококками капсульного патогенного (вирулентного) штамма мыши погибают. Капсульные пневмококки размножаются. |
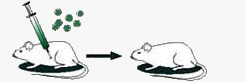 | При инфицировании пневмококками бескапсульного непатогенного штамма животные не погибают. Бескапсульные пневмококки не размножаются. |
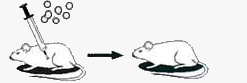 | После заражения пневмококками капсульного патогенного (вирулентного) штамма, предварительно убитыми нагреванием, мыши не погибают. Пневмококки не размножаются. |
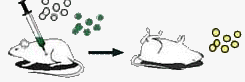 | При введении мышам одновременно бескапсульных невирулентных пневмококков и капсульных, убитых нагреванием, животные погибают. Капсульные пневмококки размножаются. |
Трансформирующим фактором оказалась ДНК. 2. 1952г. Эксперимент Альфреда Херши и Марты Чейз.Фаги (бактериофаги) - это вирусы, размножающиеся в бактериях.E. сoli - кишечная палочка (эубактерия). Суть опыта: фаги, у которых белковая оболочка была мечена радиоактивной серой (S35), а ДНК - радиоактивным фосфором (Р32), инкубировали с бактериями. Затем бактерии отмывали. В смывных водах не обнаруживали Р32, а в бактериях - S35 Следовательно, внутрь попала только ДНК. Через несколько минут из бактерии выходили десятки полноценных фагов, содержащих и белковую оболочку, и ДНК.
| Эксперимент А.Херши и М.Чейз | |
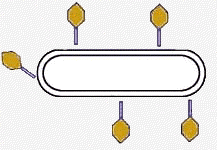 | Фаги, у которых белковая оболочка была мечена радиоактивной серой (S35), а ДНК - радиоактивным фосфором (Р32), инкубировали с бактериями. |
 | Затем бактерии отмывали. В смывных водах не обнаруживали Р32, а в бактериях - S35. В то же время большая часть Р32 оказалась внутри бактериальных клеток, а вся S35 - в окружающей среде. |
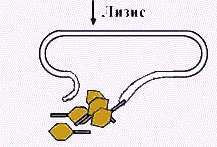 | Следовательно, внутрь попала только ДНК. Через несколько минут из бактерии выходили десятки полноценных фагов, содержащих немеченую белковую оболочку, и ДНК, меченую Р32. |
| Именно ДНК выполняет генетическую функцию - несет информацию как о создании новых копий ДНК, так и о синтезе фаговых белков | |
| Опыты Френкеля-Конрата | |
 | Френкель-Конрат работал с вирусом табачной мозаики (ВТМ). В этом вирусе содержится РНК, а не ДНК. РНК покрыта белковой облочкой. Вирус можно "разобрать" и "собрать" снова, но уже так, что РНК вируса одного штамма будет окружена белковой оболочкой вируса другого штамма. |
 | Разные штаммы вируса вызывают разную картину поражения листьев табака. После смены белковой оболочки "переодетые" вирусы вызывали картину поражения, характерную для того штамма, чья РНК была покрыта чужим белком (на рисунке - штамм 1). Позднее из пораженных растений выделяли вирусы штамма 1. |
| Не только ДНК, но и РНК может служить носителем генетической информации. | |
| Хронология открытий, подготовивших создание Уотсоном и Криком модели двойной спирали ДНК 1868г. | Обнаружен нуклеин. Современное название - хроматин. Фридрих Мишер |
| 1889г. | Нуклеин разделен на нуклеиновую кислоту и белок. Появился термин "нуклеиновая кислота". Рихард Альтман |
| 1900г. | Все азотистые основания были описаны химиками. |
| 1909г. | В нуклеиновых кислотах обнаружены фосфорная кислота и рибоза. Левин |
| 1930г. | Найдена дезоксирибоза. Левин |
| 1938г. | Рентгеноструктурный анализ показал, что расстояние между нуклеотидами в ДНК 3,4 Å. При этом азотистые основания уложены стопками. Уильям Астбюри, Флорин Белл |
| 1947г. | С помощью прямого и обратного титрования установлено, что в ДНК есть водородные связи между группами N-H и C=O. Гулланд |
| 1953г. | С помощью кислотного гидролиза ДНК с последующей хроматографией и количественным анализом установлены закономерности: А/Т=1; Г/Ц=1; (Г+Ц)/(А+Т)=К - коэффициент специфичности, постоянен для каждого вида. Эрвин Чаргафф |