Файл: Курс лекций, нгу 2000. Лекция Что такое молекулярная биология. Доказательства роли нуклеиновых кислот.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 11.01.2024
Просмотров: 158
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
канцерогены - это вещества, повышающие частоту возникновения рака.
Но в экспериментах с канцерогенами не все животные заболевали.
2. Генетическая теория.
Появилась в 30-х годах. У лабораторных мышей известны высоко- и низкораковые лабораторные линии
3. Вирусная теория.
В молоке мышей был найден "фактор молока" (вирус Битнера).
Объединение всех этих теорий произошло в 50-х годах. Отечественный ученый Лев Зильбер высказал гипотезу, что причиной рака может быть вирус, который становится геном.
Обратная транскрипция
Определение: обратная транскрипция - это синтез ДНК по матрице РНК.
Обратную транскрипцию обнаружили в 1970 г. Темин, Балтимор, Дульбеко, работавшие с вирусом саркомы Рауса (ВСР). Этот вирус вызывает саркому у кур. Это онкорнавирус (oncoRNA) - относится к ретровирусам.
Определение: ретровирусы - это РНК-содержащие вирусы, в жизненный цикл которых входит стадия образования ДНК обратной транскриптазой и внедрение ее в геном клетки хозяина в форме провируса.
Предпочтительного места внедрения провируса в геном нет. Это позволяет отнести его к мобильным генетическим элементам.
В состав ретровируса входит две идентичные молекулы РНК. На 5'-конце имеется Сap, на 3'-конце - поли А-хвост. Фермент обратную транскриптазу вирус "носит" c собой.
Геном ретровируса содержит 4 гена:
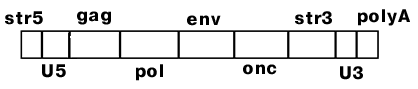
gag - белок нуклеоида,
pol - обратная транскриптаза,
env - белок капсида (оболочки),
onc - онкоген, ответственный за злокачественную трансформацию клетки.
str5 = str3 - (short terminal repeat) короткий концевой повтор;
U5, U3 - уникальные последовательности (U5 - 80 н., U3 - 200 н.);
PB (primer binding site) - участок связывания затравки.
На РВ садится (за счет комплементарности) tРНК и служит затравкой для синтеза ДНК.
Синтезируется небольшой кусок ДНК.
Обратная транскриптаза, обладая еще и активностью РНК-азы Н, удаляет РНК в гибриде с ДНК, а за счет идентичности str3 и str5 этот одноцепочечный участок ДНК взаимодействует с 3'-концом второй молекулы РНК, которая служит матрицей для продолжения синтеза цепи ДНК.
Затем РНК-матрица уничтожается и по образовавшейся цепи ДНК строится комплементарная.
Образованная молекула ДНК длиннее РНК. Она содержит LTR (U3 str 3(5) U5). В форме провируса она находится в геноме клетки хозяина. При митозе и мейозе передается дочерним клеткам и потомкам.
Для экспрессии вирусных генов нужен толчок: канцерогены, изменения метаболизма в клетке хозяина, стресс.
Большинство изученных вирусных онкогенов кодируют протеинкиназу, фермент, который фосфорилирует белки. Как правило - это тирозиновая протеинкиназа. В клетке есть собственные протеинкиназы, в том числе и тирозиновая, но гораздо более активны сериновая и треониновая. Гены, кодирующие клеточные протеинкиназы, обозначают oncc, вирусные - oncv. Oncc - клеточные гены, работающие в дифференцированных клетках. Oncc имеют интроны, oncv - не имеют. Oncv либо добавляет тирозиновую протеинкиназу - и сказывается дозовый эффект гена тирозиновой протеинкиназы, либо, по сравнению с клеткой, не имеющей oncv, клетка, его имеющая, фосфорилирует тирозин, а не серин или треонин, как обычно, то есть происходит смена мишени.
В первую очередь это касается белков, присутствующих в клетке в большом количестве. Это белки цитоскелета (нарушение адгезии), мембранные белки (нарушение контактного торможения), гистоны (нарушение регуляции, компактизации, облегчение репликации ДНК).
Ретровирусы скорее всего возникли в результате внедрения мобильных элементов в непосредственной близости от oncc генов. В дальнейшем oncc превратился в oncv, а клеточная полимераза - в обратную транскриптазу. Вирус начал самостоятельную жизнь. Стадия провируса говорит о его клеточном происхождении.
В медицине рак - это злокачественная опухоль только эпителиальных тканей.
Метастазы - возникающие опухоли в районе удаления от исходной опухоли.
Рак - болезнь генома.
Одним из путей активации oncc является такая перестройка генома, в результате которой рядом с онкогеном появляется новый регуляторный элемент, обеспечивающий его более активную транскрипцию.
Другой путь - структурная мутация в протоонкогене, т.е. нормальном клеточном гене, способном превратиться в онкоген.
Существуют антионкогены, или гены-супрессоры опухолей, подавление активности которых приводит к развитию опухолей.
Природа белковых продуктов онкогенов и антионкогенов чрезвычайно разнообразна. К онкогенам относят некоторые гены белков - факторов роста, а также гены рецепторов факторов роста. Перепроизводство факторов роста или нарушение структуры их рецепторов может привести к более частому делению клеток. Изменения в генах, кодирующих белки - передатчики сигналов от рецепторов к ядру клетки, в основном, протеинкиназы различной специфичности, а также изменения экспрессии генов, ответственных за белковые факторы транскрипции, могут превратить нормальную клетку в раковую.
Подавление активности генов, ответственных за рост и размножение клеток, осуществляется белковыми продуктами генов - супрессоров опухолей. Так, ключевая роль в разрешении на переход из одной фазы клеточного цикла в другую принадлежит белкам - циклинам. Только находясь в комплексе с циклинами, циклинзависимые протеинкиназы способны фосфорилировать белки мишени, необходимые для перехода в следующую фазу клеточного цикла.
Специальные белки сканируют ДНК перед репликацией на предмет выявления нерепарированных повреждений. Если ДНК не проходит тест, то включаются системы реализации "запрограммированной смерти" - апоптоза, в результате чего разрушаются жизненно важные структуры клетки, в том числе хромосомы и цитоскелет. Апоптоз определяется большим числом генов, центральное место среди которых занимает ген, кодирующий белок с молекулярным весом 53 кДа, - ген p53. Этот ген поврежден в 50% всех опухолей человека. Когда он выведен из строя, клетки с поврежденной (мутантной) ДНК перестают выбраковываться и в них происходит накопление новых мутаций, которые могут затрагивать как протоонкогены, так и гены-супрессоры опухолей.
Как правило, рак развивается у людей пожилого и старого возраста. Это связано с тем, что мутации возникают случайно - и вероятность накопления в клетке нужного для злокачественного превращения набора измененных генов увеличивается с годами. Посчитано, что в среднем в клетке человека должно накопиться 10 независимых мутаций, касающихся онкогенов и генов - супрессоров опухолей.
Молекулярные основы канцерогенеза
Признаки трансформированной клетки
1. Неконтролируемое деление.
Искажен клеточный цикл. Продолжителен S-период. Стадия G2 сведена к минимуму. Клетка вступает в митоз неготовой.
Последствия: нарушения при расхождении хромосом.
Высокая потребность в энергии. При этом в элокачественных клетках гликолиз (идущий без кислорода) превалирует над окислительным фосфорилированием.
2. Клетки перестают узнавать друг друга.
Происходит утрата контактного торможения. Это связано с изменением мембранных белков - белков-рецепторов и пр.
Нарушается адгезия (прилипание к поверхности).
3. Раковые клетки дедифференцированы.
Теории рака
До 70-х годов существовало три теории рака:
1. Канцерогенная теория.
Известны професиональные раковые заболевания: рак кожи у трубочистов, рак губы у кровельщиков и пр.
Бензпирен - первый описанный канцероген.
Определение: канцерогены - это вещества, повышающие частоту возникновения рака.
Но в экспериментах с канцерогенами не все животные заболевали.
2. Генетическая теория.
Появилась в 30-х годах. У лабораторных мышей известны высоко- и низкораковые лабораторные линии
3. Вирусная теория.
В молоке мышей был найден "фактор молока" (вирус Битнера).
Объединение всех этих теорий произошло в 50-х годах. Отечественный ученый Лев Зильбер высказал гипотезу, что причиной рака может быть вирус, который становится геном.
Обратная транскрипция
Определение: обратная транскрипция - это синтез ДНК по матрице РНК.
Обратную транскрипцию обнаружили в 1970 г. Темин, Балтимор, Дульбеко, работавшие с вирусом саркомы Рауса (ВСР). Этот вирус вызывает саркому у кур. Это онкорнавирус (oncoRNA) - относится к ретровирусам.
Определение: ретровирусы - это РНК-содержащие вирусы, в жизненный цикл которых входит стадия образования ДНК обратной транскриптазой и внедрение ее в геном клетки хозяина в форме провируса.
Предпочтительного места внедрения провируса в геном нет. Это позволяет отнести его к мобильным генетическим элементам.
В состав ретровируса входит две идентичные молекулы РНК. На 5'-конце имеется Сap, на 3'-конце - поли А-хвост. Фермент обратную транскриптазу вирус "носит" c собой.
Геном ретровируса содержит 4 гена:
gag - белок нуклеоида,
pol - обратная транскриптаза,
env - белок капсида (оболочки),
onc - онкоген, ответственный за злокачественную трансформацию клетки.
str5 = str3 - (short terminal repeat) короткий концевой повтор;
U5, U3 - уникальные последовательности (U5 - 80 н., U3 - 200 н.);
PB (primer binding site) - участок связывания затравки.
На РВ садится (за счет комплементарности) tРНК и служит затравкой для синтеза ДНК.
Синтезируется небольшой кусок ДНК.
Обратная транскриптаза, обладая еще и активностью РНК-азы Н, удаляет РНК в гибриде с ДНК, а за счет идентичности str3 и str5 этот одноцепочечный участок ДНК взаимодействует с 3'-концом второй молекулы РНК, которая служит матрицей для продолжения синтеза цепи ДНК.
Затем РНК-матрица уничтожается и по образовавшейся цепи ДНК строится комплементарная.
Образованная молекула ДНК длиннее РНК. Она содержит LTR (U3 str 3(5) U5). В форме провируса она находится в геноме клетки хозяина. При митозе и мейозе передается дочерним клеткам и потомкам.
Для экспрессии вирусных генов нужен толчок: канцерогены, изменения метаболизма в клетке хозяина, стресс.
Большинство изученных вирусных онкогенов кодируют протеинкиназу, фермент, который фосфорилирует белки. Как правило - это тирозиновая протеинкиназа. В клетке есть собственные протеинкиназы, в том числе и тирозиновая, но гораздо более активны сериновая и треониновая. Гены, кодирующие клеточные протеинкиназы, обозначают oncc, вирусные - oncv. Oncc - клеточные гены, работающие в дифференцированных клетках. Oncc имеют интроны, oncv - не имеют. Oncv либо добавляет тирозиновую протеинкиназу - и сказывается дозовый эффект гена тирозиновой протеинкиназы, либо, по сравнению с клеткой, не имеющей oncv, клетка, его имеющая, фосфорилирует тирозин, а не серин или треонин, как обычно, то есть происходит смена мишени.
В первую очередь это касается белков, присутствующих в клетке в большом количестве. Это белки цитоскелета (нарушение адгезии), мембранные белки (нарушение контактного торможения), гистоны (нарушение регуляции, компактизации, облегчение репликации ДНК).
Ретровирусы скорее всего возникли в результате внедрения мобильных элементов в непосредственной близости от oncc генов. В дальнейшем oncc превратился в oncv, а клеточная полимераза - в обратную транскриптазу. Вирус начал самостоятельную жизнь. Стадия провируса говорит о его клеточном происхождении.
В медицине рак - это злокачественная опухоль только эпителиальных тканей.
Метастазы - возникающие опухоли в районе удаления от исходной опухоли.
Рак - болезнь генома.
Одним из путей активации oncc является такая перестройка генома, в результате которой рядом с онкогеном появляется новый регуляторный элемент, обеспечивающий его более активную транскрипцию.
Другой путь - структурная мутация в протоонкогене, т.е. нормальном клеточном гене, способном превратиться в онкоген.
Существуют антионкогены, или гены-супрессоры опухолей, подавление активности которых приводит к развитию опухолей.
Природа белковых продуктов онкогенов и антионкогенов чрезвычайно разнообразна. К онкогенам относят некоторые гены белков - факторов роста, а также гены рецепторов факторов роста. Перепроизводство факторов роста или нарушение структуры их рецепторов может привести к более частому делению клеток. Изменения в генах, кодирующих белки - передатчики сигналов от рецепторов к ядру клетки, в основном, протеинкиназы различной специфичности, а также изменения экспрессии генов, ответственных за белковые факторы транскрипции, могут превратить нормальную клетку в раковую.
Подавление активности генов, ответственных за рост и размножение клеток, осуществляется белковыми продуктами генов - супрессоров опухолей. Так, ключевая роль в разрешении на переход из одной фазы клеточного цикла в другую принадлежит белкам - циклинам. Только находясь в комплексе с циклинами, циклинзависимые протеинкиназы способны фосфорилировать белки мишени, необходимые для перехода в следующую фазу клеточного цикла.
Специальные белки сканируют ДНК перед репликацией на предмет выявления нерепарированных повреждений. Если ДНК не проходит тест, то включаются системы реализации "запрограммированной смерти" - апоптоза, в результате чего разрушаются жизненно важные структуры клетки, в том числе хромосомы и цитоскелет. Апоптоз определяется большим числом генов, центральное место среди которых занимает ген, кодирующий белок с молекулярным весом 53 кДа, - ген p53. Этот ген поврежден в 50% всех опухолей человека. Когда он выведен из строя, клетки с поврежденной (мутантной) ДНК перестают выбраковываться и в них происходит накопление новых мутаций, которые могут затрагивать как протоонкогены, так и гены-супрессоры опухолей.
Как правило, рак развивается у людей пожилого и старого возраста. Это связано с тем, что мутации возникают случайно - и вероятность накопления в клетке нужного для злокачественного превращения набора измененных генов увеличивается с годами. Посчитано, что в среднем в клетке человека должно накопиться 10 независимых мутаций, касающихся онкогенов и генов - супрессоров опухолей.
Гипотезы возникновения жизни
Панспермия - жизнь витает в космосе и разносится по планетам.
Жизнь зародилась абиогенно или нет?
Биогенез - живое только от живого.
Абиогенез - живое от неживого.
Луи Пастеру принадлежит первое прямое доказательство происхождения живого только от живого. В 1862 году он получил премию Французской академии наук за эту работу.
Суть опыта: в колбе с изогнутой трубкой находился прокипяченный сенный настой. В течение нескольких недель он стоял совершенно прозрачный. Как только колбу наклонили (сквозь трубку в колбу попали микроорганизмы) - настой забродил.
Эксперимент правильный. Вывод - живое только от живого.
Авторитет Пастера был столь велик, что к теории абиогенеза пришли лишь через 60 лет.
В 1924 году Александр Опарин высказал предположение, что
4 млд. лет назад жизнь могла возникнуть абиогенно, в силу тех условий, которые существовали тогда на Земле.
Джон Холдейн рассчитал, какие условия и как долго должны были существовать, чтобы зародилась жизнь, каковы необходимые источники энергии для зарождения жизни.
Теория биопоэза
Джон Бернал создал теорию биопоэза, включающую три стадии.
1. Образование биомономеров.
2. Образование биополимеров и их эволюция. Образование систем с обратной связью.
3. Образование мембранных структур и пробионтов (первых клеток).
Экспериментальное доказательство первой стадии - опыты Стенли Миллера.
Суть опыта: в колбе находилась смесь газов (H2, N2, NH3, CH4, CO, CO2) при температуре
100
Но в экспериментах с канцерогенами не все животные заболевали.
2. Генетическая теория.
Появилась в 30-х годах. У лабораторных мышей известны высоко- и низкораковые лабораторные линии
3. Вирусная теория.
В молоке мышей был найден "фактор молока" (вирус Битнера).
Объединение всех этих теорий произошло в 50-х годах. Отечественный ученый Лев Зильбер высказал гипотезу, что причиной рака может быть вирус, который становится геном.
Обратная транскрипция
Определение: обратная транскрипция - это синтез ДНК по матрице РНК.
Обратную транскрипцию обнаружили в 1970 г. Темин, Балтимор, Дульбеко, работавшие с вирусом саркомы Рауса (ВСР). Этот вирус вызывает саркому у кур. Это онкорнавирус (oncoRNA) - относится к ретровирусам.
Определение: ретровирусы - это РНК-содержащие вирусы, в жизненный цикл которых входит стадия образования ДНК обратной транскриптазой и внедрение ее в геном клетки хозяина в форме провируса.
Предпочтительного места внедрения провируса в геном нет. Это позволяет отнести его к мобильным генетическим элементам.
В состав ретровируса входит две идентичные молекулы РНК. На 5'-конце имеется Сap, на 3'-конце - поли А-хвост. Фермент обратную транскриптазу вирус "носит" c собой.
| Геном ретровируса содержит 4 гена: | |
gag - белок нуклеоида,
pol - обратная транскриптаза,
env - белок капсида (оболочки),
onc - онкоген, ответственный за злокачественную трансформацию клетки.
str5 = str3 - (short terminal repeat) короткий концевой повтор;
U5, U3 - уникальные последовательности (U5 - 80 н., U3 - 200 н.);
PB (primer binding site) - участок связывания затравки.
| | На РВ садится (за счет комплементарности) tРНК и служит затравкой для синтеза ДНК. Синтезируется небольшой кусок ДНК. |
Обратная транскриптаза, обладая еще и активностью РНК-азы Н, удаляет РНК в гибриде с ДНК, а за счет идентичности str3 и str5 этот одноцепочечный участок ДНК взаимодействует с 3'-концом второй молекулы РНК, которая служит матрицей для продолжения синтеза цепи ДНК.
Затем РНК-матрица уничтожается и по образовавшейся цепи ДНК строится комплементарная.
Образованная молекула ДНК длиннее РНК. Она содержит LTR (U3 str 3(5) U5). В форме провируса она находится в геноме клетки хозяина. При митозе и мейозе передается дочерним клеткам и потомкам.
Для экспрессии вирусных генов нужен толчок: канцерогены, изменения метаболизма в клетке хозяина, стресс.
Большинство изученных вирусных онкогенов кодируют протеинкиназу, фермент, который фосфорилирует белки. Как правило - это тирозиновая протеинкиназа. В клетке есть собственные протеинкиназы, в том числе и тирозиновая, но гораздо более активны сериновая и треониновая. Гены, кодирующие клеточные протеинкиназы, обозначают oncc, вирусные - oncv. Oncc - клеточные гены, работающие в дифференцированных клетках. Oncc имеют интроны, oncv - не имеют. Oncv либо добавляет тирозиновую протеинкиназу - и сказывается дозовый эффект гена тирозиновой протеинкиназы, либо, по сравнению с клеткой, не имеющей oncv, клетка, его имеющая, фосфорилирует тирозин, а не серин или треонин, как обычно, то есть происходит смена мишени.
В первую очередь это касается белков, присутствующих в клетке в большом количестве. Это белки цитоскелета (нарушение адгезии), мембранные белки (нарушение контактного торможения), гистоны (нарушение регуляции, компактизации, облегчение репликации ДНК).
Ретровирусы скорее всего возникли в результате внедрения мобильных элементов в непосредственной близости от oncc генов. В дальнейшем oncc превратился в oncv, а клеточная полимераза - в обратную транскриптазу. Вирус начал самостоятельную жизнь. Стадия провируса говорит о его клеточном происхождении.
В медицине рак - это злокачественная опухоль только эпителиальных тканей.
Метастазы - возникающие опухоли в районе удаления от исходной опухоли.
Рак - болезнь генома.
Одним из путей активации oncc является такая перестройка генома, в результате которой рядом с онкогеном появляется новый регуляторный элемент, обеспечивающий его более активную транскрипцию.
Другой путь - структурная мутация в протоонкогене, т.е. нормальном клеточном гене, способном превратиться в онкоген.
Существуют антионкогены, или гены-супрессоры опухолей, подавление активности которых приводит к развитию опухолей.
Природа белковых продуктов онкогенов и антионкогенов чрезвычайно разнообразна. К онкогенам относят некоторые гены белков - факторов роста, а также гены рецепторов факторов роста. Перепроизводство факторов роста или нарушение структуры их рецепторов может привести к более частому делению клеток. Изменения в генах, кодирующих белки - передатчики сигналов от рецепторов к ядру клетки, в основном, протеинкиназы различной специфичности, а также изменения экспрессии генов, ответственных за белковые факторы транскрипции, могут превратить нормальную клетку в раковую.
Подавление активности генов, ответственных за рост и размножение клеток, осуществляется белковыми продуктами генов - супрессоров опухолей. Так, ключевая роль в разрешении на переход из одной фазы клеточного цикла в другую принадлежит белкам - циклинам. Только находясь в комплексе с циклинами, циклинзависимые протеинкиназы способны фосфорилировать белки мишени, необходимые для перехода в следующую фазу клеточного цикла.
Специальные белки сканируют ДНК перед репликацией на предмет выявления нерепарированных повреждений. Если ДНК не проходит тест, то включаются системы реализации "запрограммированной смерти" - апоптоза, в результате чего разрушаются жизненно важные структуры клетки, в том числе хромосомы и цитоскелет. Апоптоз определяется большим числом генов, центральное место среди которых занимает ген, кодирующий белок с молекулярным весом 53 кДа, - ген p53. Этот ген поврежден в 50% всех опухолей человека. Когда он выведен из строя, клетки с поврежденной (мутантной) ДНК перестают выбраковываться и в них происходит накопление новых мутаций, которые могут затрагивать как протоонкогены, так и гены-супрессоры опухолей.
Как правило, рак развивается у людей пожилого и старого возраста. Это связано с тем, что мутации возникают случайно - и вероятность накопления в клетке нужного для злокачественного превращения набора измененных генов увеличивается с годами. Посчитано, что в среднем в клетке человека должно накопиться 10 независимых мутаций, касающихся онкогенов и генов - супрессоров опухолей.
Молекулярные основы канцерогенеза
Признаки трансформированной клетки
1. Неконтролируемое деление.
Искажен клеточный цикл. Продолжителен S-период. Стадия G2 сведена к минимуму. Клетка вступает в митоз неготовой.
Последствия: нарушения при расхождении хромосом.
Высокая потребность в энергии. При этом в элокачественных клетках гликолиз (идущий без кислорода) превалирует над окислительным фосфорилированием.
2. Клетки перестают узнавать друг друга.
Происходит утрата контактного торможения. Это связано с изменением мембранных белков - белков-рецепторов и пр.
Нарушается адгезия (прилипание к поверхности).
3. Раковые клетки дедифференцированы.
Теории рака
До 70-х годов существовало три теории рака:
1. Канцерогенная теория.
Известны професиональные раковые заболевания: рак кожи у трубочистов, рак губы у кровельщиков и пр.
Бензпирен - первый описанный канцероген.
Определение: канцерогены - это вещества, повышающие частоту возникновения рака.
Но в экспериментах с канцерогенами не все животные заболевали.
2. Генетическая теория.
Появилась в 30-х годах. У лабораторных мышей известны высоко- и низкораковые лабораторные линии
3. Вирусная теория.
В молоке мышей был найден "фактор молока" (вирус Битнера).
Объединение всех этих теорий произошло в 50-х годах. Отечественный ученый Лев Зильбер высказал гипотезу, что причиной рака может быть вирус, который становится геном.
Обратная транскрипция
Определение: обратная транскрипция - это синтез ДНК по матрице РНК.
Обратную транскрипцию обнаружили в 1970 г. Темин, Балтимор, Дульбеко, работавшие с вирусом саркомы Рауса (ВСР). Этот вирус вызывает саркому у кур. Это онкорнавирус (oncoRNA) - относится к ретровирусам.
Определение: ретровирусы - это РНК-содержащие вирусы, в жизненный цикл которых входит стадия образования ДНК обратной транскриптазой и внедрение ее в геном клетки хозяина в форме провируса.
Предпочтительного места внедрения провируса в геном нет. Это позволяет отнести его к мобильным генетическим элементам.
В состав ретровируса входит две идентичные молекулы РНК. На 5'-конце имеется Сap, на 3'-конце - поли А-хвост. Фермент обратную транскриптазу вирус "носит" c собой.
| Геном ретровируса содержит 4 гена: | |
gag - белок нуклеоида,
pol - обратная транскриптаза,
env - белок капсида (оболочки),
onc - онкоген, ответственный за злокачественную трансформацию клетки.
str5 = str3 - (short terminal repeat) короткий концевой повтор;
U5, U3 - уникальные последовательности (U5 - 80 н., U3 - 200 н.);
PB (primer binding site) - участок связывания затравки.
| | На РВ садится (за счет комплементарности) tРНК и служит затравкой для синтеза ДНК. Синтезируется небольшой кусок ДНК. |
Обратная транскриптаза, обладая еще и активностью РНК-азы Н, удаляет РНК в гибриде с ДНК, а за счет идентичности str3 и str5 этот одноцепочечный участок ДНК взаимодействует с 3'-концом второй молекулы РНК, которая служит матрицей для продолжения синтеза цепи ДНК.
Затем РНК-матрица уничтожается и по образовавшейся цепи ДНК строится комплементарная.
Образованная молекула ДНК длиннее РНК. Она содержит LTR (U3 str 3(5) U5). В форме провируса она находится в геноме клетки хозяина. При митозе и мейозе передается дочерним клеткам и потомкам.
Для экспрессии вирусных генов нужен толчок: канцерогены, изменения метаболизма в клетке хозяина, стресс.
Большинство изученных вирусных онкогенов кодируют протеинкиназу, фермент, который фосфорилирует белки. Как правило - это тирозиновая протеинкиназа. В клетке есть собственные протеинкиназы, в том числе и тирозиновая, но гораздо более активны сериновая и треониновая. Гены, кодирующие клеточные протеинкиназы, обозначают oncc, вирусные - oncv. Oncc - клеточные гены, работающие в дифференцированных клетках. Oncc имеют интроны, oncv - не имеют. Oncv либо добавляет тирозиновую протеинкиназу - и сказывается дозовый эффект гена тирозиновой протеинкиназы, либо, по сравнению с клеткой, не имеющей oncv, клетка, его имеющая, фосфорилирует тирозин, а не серин или треонин, как обычно, то есть происходит смена мишени.
В первую очередь это касается белков, присутствующих в клетке в большом количестве. Это белки цитоскелета (нарушение адгезии), мембранные белки (нарушение контактного торможения), гистоны (нарушение регуляции, компактизации, облегчение репликации ДНК).
Ретровирусы скорее всего возникли в результате внедрения мобильных элементов в непосредственной близости от oncc генов. В дальнейшем oncc превратился в oncv, а клеточная полимераза - в обратную транскриптазу. Вирус начал самостоятельную жизнь. Стадия провируса говорит о его клеточном происхождении.
В медицине рак - это злокачественная опухоль только эпителиальных тканей.
Метастазы - возникающие опухоли в районе удаления от исходной опухоли.
Рак - болезнь генома.
Одним из путей активации oncc является такая перестройка генома, в результате которой рядом с онкогеном появляется новый регуляторный элемент, обеспечивающий его более активную транскрипцию.
Другой путь - структурная мутация в протоонкогене, т.е. нормальном клеточном гене, способном превратиться в онкоген.
Существуют антионкогены, или гены-супрессоры опухолей, подавление активности которых приводит к развитию опухолей.
Природа белковых продуктов онкогенов и антионкогенов чрезвычайно разнообразна. К онкогенам относят некоторые гены белков - факторов роста, а также гены рецепторов факторов роста. Перепроизводство факторов роста или нарушение структуры их рецепторов может привести к более частому делению клеток. Изменения в генах, кодирующих белки - передатчики сигналов от рецепторов к ядру клетки, в основном, протеинкиназы различной специфичности, а также изменения экспрессии генов, ответственных за белковые факторы транскрипции, могут превратить нормальную клетку в раковую.
Подавление активности генов, ответственных за рост и размножение клеток, осуществляется белковыми продуктами генов - супрессоров опухолей. Так, ключевая роль в разрешении на переход из одной фазы клеточного цикла в другую принадлежит белкам - циклинам. Только находясь в комплексе с циклинами, циклинзависимые протеинкиназы способны фосфорилировать белки мишени, необходимые для перехода в следующую фазу клеточного цикла.
Специальные белки сканируют ДНК перед репликацией на предмет выявления нерепарированных повреждений. Если ДНК не проходит тест, то включаются системы реализации "запрограммированной смерти" - апоптоза, в результате чего разрушаются жизненно важные структуры клетки, в том числе хромосомы и цитоскелет. Апоптоз определяется большим числом генов, центральное место среди которых занимает ген, кодирующий белок с молекулярным весом 53 кДа, - ген p53. Этот ген поврежден в 50% всех опухолей человека. Когда он выведен из строя, клетки с поврежденной (мутантной) ДНК перестают выбраковываться и в них происходит накопление новых мутаций, которые могут затрагивать как протоонкогены, так и гены-супрессоры опухолей.
Как правило, рак развивается у людей пожилого и старого возраста. Это связано с тем, что мутации возникают случайно - и вероятность накопления в клетке нужного для злокачественного превращения набора измененных генов увеличивается с годами. Посчитано, что в среднем в клетке человека должно накопиться 10 независимых мутаций, касающихся онкогенов и генов - супрессоров опухолей.
Гипотезы возникновения жизни
Панспермия - жизнь витает в космосе и разносится по планетам.
Жизнь зародилась абиогенно или нет?
Биогенез - живое только от живого.
Абиогенез - живое от неживого.
Луи Пастеру принадлежит первое прямое доказательство происхождения живого только от живого. В 1862 году он получил премию Французской академии наук за эту работу.
Суть опыта: в колбе с изогнутой трубкой находился прокипяченный сенный настой. В течение нескольких недель он стоял совершенно прозрачный. Как только колбу наклонили (сквозь трубку в колбу попали микроорганизмы) - настой забродил.
Эксперимент правильный. Вывод - живое только от живого.
Авторитет Пастера был столь велик, что к теории абиогенеза пришли лишь через 60 лет.
В 1924 году Александр Опарин высказал предположение, что
0C. Кипящая вода служила источником водяного пара, а с помощью обратного холодильника поддерживалась циркуляция газовой смеси через сосуд. Давали искровой разряд в 60 тыс. вольт, что энергетически эквивалентно 50-и млн. лет на примитивной Земле. Результат был ошеломляющий: в колбе появились HCN, HCHO, HCOOH, несколько аминокислот, несколько азотистых оснований жирные кислоты, псирты, моносахара. Эксперимент повторяли много раз. Неперменное условие успеха - отсутствие в колбе свободного кислорода. В зависимости от pH раствора и соотношения газов были получены разные наборы соединений. Если была H3PO4, то образовывались даже нуклеотиды, а это уже гетерополимеры.
Таким образом была доказана первая стадия возникновения жизни. 4 млрд. лет тому назад с неизбежностью должны были возникнуть биомономеры.
Первичная атмосфера образующейся Земли кислород содержала, но он весь пошел на окисление. Свободного кислорода не было. Таким образом, возникновение биомономеров и биополимеров происходило во вторичной бескилородной среде.
У стадии 3 в принципе есть доказательства. Самая сложная и неочевидная - стадия 2.
2 стадия биопоэза.
Помимо 4-х основных классов биополимеров, могли образовываться и не дошедшие до нас гетерополимеры. Видимо, эволюция химических соединений шла по принципу минимума свободной энергии.
Остановимся пока на белках и нуклеиновых кислотах.
Из разных комплексов белок-нуклеиновая кислота рассмотрим только те, в которых
нуклеиновая кислота сохраняется благодаря защите белком от ультрафиолетового излучения.
Накопим такие комплексы. Из их множества рассмотрим те, в которых белки способствуют увеличению количества защищенной нуклеиновой кислоты. То есть эти белки - ферменты. Из этих комплексов рассмотрим те, где нуклеиновые кислоты, количество которых возрастает под действием белков, способствуют увеличению количества белков благодаря, например, прямому кодированию. Возникают системы с обратной связью. Такие системы обладают некоторыми признаками живого.
Другой вариант.
Первыми молекулами были РНК.
Они имеют третичную структуру и обладают каталитической активностью. Позже появились белки, поддерживающие "выгодные" конформации РНК и защищающие их от расщепления. Уже потом возникает ДНК, как более надежный хранитель генетической информации. Она имеет две цепи, что обеспечивает репарацию, репликация осуществляется за один шаг. Отсутствие ОН-группы в 2'-положении пентозы делает ДНК устойчивой в слабощелочных условиях, губительных для РНК.
Стадия 3.
Представим, что лужа покрыта жирной пленкой, а под ней - белки. Если оторвать каплю, то могут получиться пузырьки, содержащие нуклеопротеидные системы с обратной связью. Когда они падают на поверхность водоема, то покрываются вторым липидно-белковым слоем - и образуется современная биологическая мембрана. В мембранной капле диффузия уже не очень существенна.
Далее образуются пробионты - первые организмы, имеющие мембрану.
Эволюция пробиотов
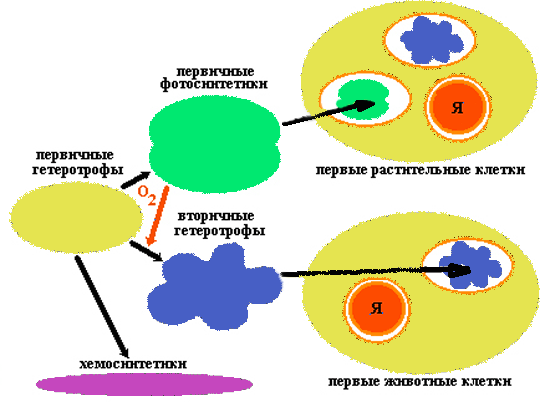 |
Пробионты были первичными гетеротрофами. Они получали энергию при расщеплении органических веществ абиогенного происхождения, в изобилии имевшихся в окружающей среде. Примером древнего способа обмена веществ, дошедшего до наших дней, является гликолиз - ферментативное бескислородное расщепление глюкозы.
По мере истощения запаса органического материала (а новый не образовывался из-за изменения условий на Земле) возникала жесткая конкурентная борьба за него, что ускорило процесс эволюции первичных гетеротрофов.
Исключительным событием стало возникновение бактериального фотосинтеза, освободившего клетки от зависимости от доступности органики абиогенного происхождения. Скорее всего, фотосинтез возник у анаэробных бактерий, способных к азотофиксации. Побочным продуктом фотосинтеза является кислород. Его накопление в атмосфере привело к коренному изменению хода эволюции. Появление озонового экрана защитило первичные организмы от смертельного УФ-облучения и положило конец абиогенному синтезу органики.
Первые аэробные бактерии появились благодаря приобретению аппарата окислительного фосфорилирования. Продукты брожения подвергались дальнейшему окислению до СО2 и Н2О. Аэробные (вторичные) гетеротрофы могли более эффективно, чем анаэробные (первичные) гетеротрофы, расщеплять органические вещества, образующиеся в результате фотосинтеза.
По-видимому, с ростом концентрации кислорода в атмосфере усложнялась жизнь первичных анаэробных гетеротрофов. Некоторые из них вымерли, другие нашли бескислородную среду. Примером могут служить дошедшие до наших дней метанобразующие бактерии или серные бактерии, живущие в горячих подземных источниках.
Некоторые первичные гетеротрофы пошли по пути, приведшему к образованию эукариотических клеток. Часть из них вступила в симбиоз с аэробными бактериями, способными к окислительному фосфорилированию. Поглотив вторичных гетеротрофов, первичные не расщепили их на молекулы, а сохранили в качестве энергетических станций, называемых сегодня митохондриями.
Такие симбионты дали начало царствам животных и грибов.
Другая часть первичных гетеротрофов "заключила союз" не только с аэробными гетеротрофами, но и с первичными фотосинтетиками, сохранив последних в качестве хлоропластов. Такие симбионты дали начало царству растений.
В пользу симбиотической теории образования эукариот говорят следующие факты:
- У митохондрий и хлоропластов две мембраны. Внутренняя - своя, наружняя образована клеткой-захватчиком.
- Генетический код митохондрий идеален. Универсальный генетический код имеет два существенных отличия, касающихся инициации и терминации синтеза белка.
Таким образом эукариоты отстранились от чужой генетической информации.
Кроме того, они линеаризовали свою ДНК. Митохондрии и хлоропласты имеют кольцевую ДНК, хотя не очень понятно, для чего им нужна кольцевая ДНК, и бактериальные рибосомы. Однако понятно, почему у них такая ДНК и такие рибосомы. Потому, что их предки были бактериями. Сегодня часть генов митохондриальных белков и белков хлоропластов, в том числе их РНК- и ДНК-полимераз, находятся в ядре. Вероятно, попали они туда с помощью мобильных элементов.
Все бактерии делятся на эубактерии (в том числе E.сoli) и археобактерии. Принципиальное отличие между ними в том, что гены археобактерий имеют экзон - интронное строение и сплайсинг. Эубактерии - результат эволюции ана- и аэробных гетеротрофов. Их эволюция шла в благоприятных условиях и они сменили больше поколений, избавившись от интронов. Археобактерии живут в экстремальных условиях: горячие, кислые, высокосолевые подземные воды. Эукариоты и археобактерии сохранили экзон - интронную структуру, что говорит о древнем происхождении экзонов и интронов.