Файл: Курс лекций, нгу 2000. Лекция Что такое молекулярная биология. Доказательства роли нуклеиновых кислот.doc
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 11.01.2024
Просмотров: 155
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Правило 1. 5' и 3' концы интрона очень консервативны: 5'(ГT-интрон-AГ)3' .
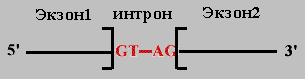
Правило 2. При сшивании копий экзонов соблюдается порядок их расположения в гене, но могут быть выброшены некоторые из них.
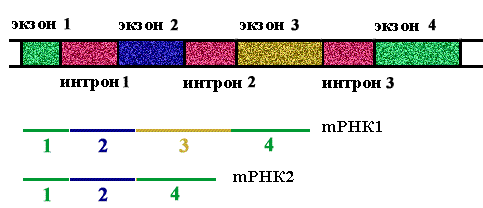
Представление об интроне, как пустой, ничего не кодирующей последовательности, неверно. Некоторые интроны кодируют ферменты-матюразы, вырезающие копии этих интронов. На вопрос, зачем эукариотическим геномам экзон - интронная структура, можно ответить только в единичных случаях. Почему эукариотические гены "разорваны", будет рассмотрено в конце курса.
Альтернативный сплайсинг mРНК кальцитонинового гена у млекопитающих (крыса)
Во всех клетках есть кальцитониновый ген, но в клетках щитовидной железы он экспрессируется в виде гормона кальцитонина, а в клетках гипофиза - нейропептида CGRP (пептида, имеющего отношение к гену кальцитонина). Ген один, а белки получаются разные в результате сплайсинга mРНК и процессинга полипептидов. В клетках других тканей этот ген не экспрессируется.
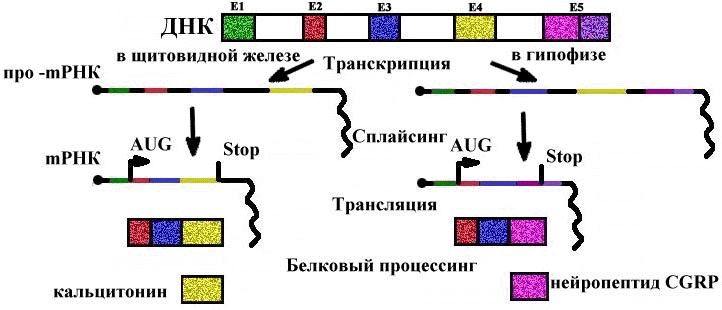
Сплайсинг осуществляется белковыми комплексами - сплайсосомами, в которых помимо ферментов, вырезающих и сшивающих участки про-mРНК, имеются белки, придающие про-mРНК нужную конформацию, и несколько sPНК. Сплайсосома непосредственно связана с ферментами, занимающимися полиаденилированием.
Автосплайсинг
Автосплайсинг открыт Томасом Чеком (США) в 1982 году.
Он работал с инфузорией Tetrаchymenа thermophyla. У этой инфузории образуется 35S про-rРНК длиной 6400 нуклеотидов. Без участия дополнительных соединений белковой природы из этой про-rРНК вырезается внутренний участок длиной в 414 нуклеотида. Два экзона сшиваются с образованием 26S rРНК. Единственное требование - определенная концентрация ионов магния. Про-rРНК имеет третичную структуру и обладает каталитичекой активностью. Впервые было показано, что каталитической активностью обладают не только белки.
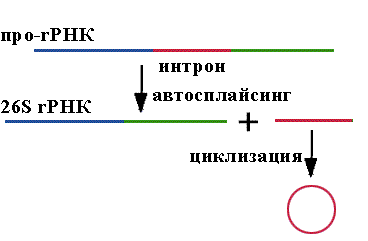
Определение: РНК-зимы - РНК с каталитической активностью.
Сегодня описано несколько десятков РНК-зимов.
Малые РНК
sРНК обнаружены в количестве 103-105 копий на клетку. Поскольку в большинстве случаев эти РНК обогащены урацилом, они называются U1, U2... Их размер от 100 до 300 нукл. Все они кодируются в ядре, но работают как в ядре (small nuclear - SN), так и в цитоплазме (small cytoplasmic - SC).
SNURPS - РНП (рибонуклеопротеидные комплексы) в ядре. snРНК входят в состав РНП, участвующих в полиаденилированиии и сплайсинге.
SCURPS - РНП в цитоплазме. Входят в состав информосом.
Малая РНК U4 присутствует в комплексах, участвующих в полиаденилировании. Если получить антитела к белкам, связывающимся с U4, то не происходит полиаденилирования и сплайсинга.
При красной волчанке (аутоимунном заболевании) вырабатываются антитела к белкам комплекса с U4.
Гистоновая mРНК не полиаденилируется потому, что sРНКU7, которая комплементарна 3'-концу гистоновой mРНК, защищает ее от полиаденилирования.
Малые РНК U1, U2, U4, U5, U6 входят в состав сплайсосомы.
Редактирование
Определение: редактирование - изменение генетической информации на уровне mРНК.
Трипаносома - одноклеточный паразит, вызывающий у человека сонную болезнь. В клетке трипаносомы есть митохондрии и множество ферментов окислительного фосфорилирования. Один из ключевых ферментов - цитохромоксидаза. Он имеет четвертичную структуру и состоит из 3-х разных субъединиц.
Человек (постоянный хозяин трипаносомы) теплокровен, поэтому трипаносоме не нужна энергия собственных митохондрий. Синтезируются только две субъединицы цитохромоксидазы.
Муха цеце (промежуточный хозяин трипаносомы) - холоднокровна. Трипаносоме нужен работающий фермент. Все три субъединицы синтезируются.
В геноме трипаносомы только два гена для двух субъединиц. В mРНК одного из них происходит разрезание и встраивание 4-х урацилов (на небольшом расстоянии друг от друга, но не подряд).
Происходит сдвиг рамки считывания и отредактированная mРНК кодирует новый полипептид - третью субъединицу цитохромоксидазы.
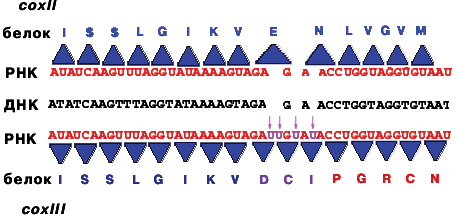
Репликация ДНК
Определение: процесс, осуществляемый комплексом ферментов и белков, выполняющих топологическую функцию, суть которого в образовании идентичных копий ДНК для передачи генетической информации в поколениях клеток и организмов, называют
репликацией ДНК.
Принципы репликации
1. Комплементарность.
2. Антипараллельность.
3. Униполярность.
4. Потребность в затравке.
5. Прерывистость.
6.Полуконсервативность.
Первые три принципа можно сформулировать в одной фразе:
Синтез каждой дочерней цепи ДНК идет комплементарно и антипараллельно матричной цепи и всегда в направлении 5' 3'.
Доказательство полуконсервативного характера репликации
Для выяснения вопроса о характере расхождения цепей по дочерним молекулам Мэтт Мезельсон и Фрэнк Сталь в 1958г. разработали метод равновесного центрифугирования в градиенте плотности CsCl. .
ДНК разделяется не по молекулярным весам, а по удельной плотности.
E. сoli выращивали на протяжении нескольких поколений на среде, содержащей тяжелый изотоп азота (N15), для того, чтобы вся ДНК была "тяжелой". Перед очередным раундом деления клетки синхронизировали. При этом в среде заменяли N15 на легкий изотоп N14 с тем, чтобы вновь синтезированные цепи были "легкими". После репликации ДНК выделяли и центрифугировали в градиенте плотности CsCl.
Полуконсервативность означает, что каждая дочерняя ДНК состоит из одной матричной цепи и одной вновь синтезированной.
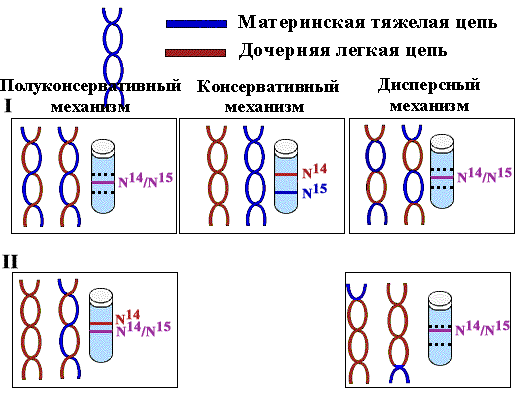
Равное распределение "тяжелых" и "легких" цепей между всеми молекулами исключало возможность консервативного способа, согласно которому одна дочерняя клетка получает материнскую ДНК, а другая - вновь синтезированную, обе цепи которой являются новыми.
Клетки второго поколения содержали как полностью "легкие" молекулы, так и "гибридные", состоящие из одной "легкой" и одной "тяжелой" цепи, аналогичные молекулам первого поколения. Этот факт исключал возможность дисперсного механизма, согластно которому куски материнской ДНК случайным образом распределяются между дочерними молекулами.
Ферментативная система синтеза ДНК in vitro
В 1956 г. Артур Корнберг наработал 100 кг биомассы E. coli и выделил 0.5 г фермента ДНК-полимеразы.
Необходимые компоненты для синтеза ДНК in vitro:
1. ДНК-матрица - образец, по которому строится новая цепь ДНК.
2. Активированные нуклеотиды (dАТФ, dГТФ, dТТФ, dЦТФ) - то, из чего строятся дочерние цепи.
3. ДНК-полимераза - то, что строит новую цепь ДНК.
4. Ионы магния - то, без чего фермент не работает.
ДНК-матрицу необходимо активировать.
Нативная двуцепочечная ДНК, не имеющая повреждений, не может эффективно использоваться в этой системе. Активировать ее можно либо денатурацией щелочью или нагреванием (1), либо обработкой экзонуклеазой III из E. сoli (2), либо внесением ников (одноцепочечных разрывов) с помощью эндонуклеаз (3).
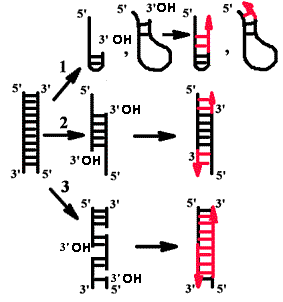
Понятие о матрице и затравке
Продукты, образуемые в ферментативной системе in vitro.
Во всех случаях матрицей для синтеза новых цепей служит одноцепочечная ДНК. Затравкой является 3'-гидроксильный конец двуцепочечной ДНК, причем он должен быть спарен с матрицей.
В том случае, если эндонуклеаза вносила ники с 3'-фосфатным концом, ДНК не являлась активированной.
Прямым доказательством того, что затравка - 3'-гидроксильный конец, является эксперимент с дидезоксинуклеозидтрифосфатом.
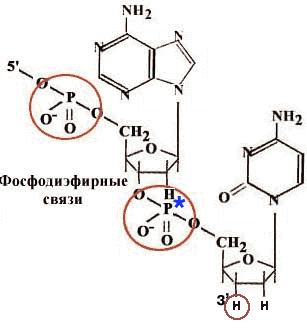 | Если такой активированный нуклеотид сделать меченым по α-фосфату, то он включается в растущую полимерную цепь и всегда обнаруживается на ее 3'-конце. Это говорит о том, что он сам включается, но дальнейший рост цепи невозможен, т.к. нет 3'-гидроксильного конца. |
Это также доказывает и униполярность репликации в направлении 5' 3'.
Строение и свойства ДНК-полимеразы Корнберга (ДНК-полимеразы I)
ДНК - полимераза Корнберга (ДНК-полимераза I) - это одна полипептидная цепь с молекулярным весом 109 тыс.
В состав полимеразы входят ионы цинка. Она абсолютно зависима от ионов магния.
Обнаружено 4 разные каталитические активности ДНК - полимеразы I:
1. Полимеризационная в направлении 5` 3`.
ДНКn + dХТФ ДНКn+1 + Ф-Ф
2. Пирофосфоролиз:
ДНКn + Ф-Ф* ДНК
n-1 + dХТФ*
Эта активность возможна при большом избытке пирофосфатов.
3. Пирофосфатный обмен:
ДНКn + dХТФ + Ф-Ф* ДНКn+ dХТФ* + Ф-Ф
Фермент работает только тогда, когда он находится на молекуле ДНК и имеет соответствующую конформацию.
4. Гидролитическая активность:
ДНКn + Н2О ДНКn-1 + dХMФ
Эта активность имеет большой биологический смысл.
Гидролитическая активность проявляется в направлении 3' 5' и 5' 3'.
Активность 3' 5' проявляется на неспаренном 3'-гидроксильном конце. Фермент возвращается при ошибке включения и "откусывает" неправильный нуклеотид.
Это корректорская функция фермента.
Все ДНК-полимеразы обладают этой активностью.