ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 01.03.2019
Просмотров: 6555
Скачиваний: 2

101
клетки, обеспечивающие гуморальный иммунный ответ. Т-лимфоциты,
дифференцируются в эффекторные Т-лимфоциты, обеспечивающие
клеточный иммунный ответ;
6) нейтрализация и уничтожение чужеродного Аг как результат
перечисленных событий.
Одновременно с активацией Т-хелперов стимулируются Т-супрессоры,
тормозящие иммунный ответ. Поэтому через определенное время в норме
иммунная реакция затихает. В организме остается иммунологическая
память: Т- и В-клетки памяти.
Деление на клеточный и гуморальный иммунный ответ условно и
определяется конечным звеном иммунного ответа: АТ или Т-эффекторы.
Кооперативный механизм действия и регуляция иммунной системы
Антиген сам является важным фактором иммунорегуляции. От
природы антигена зависит тип иммунного ответа. На полисахаридные
антигены бактерий, как правило, развивается ГИО. На бактериальные
белковые антигены — как КИО, так и ГИО. На внутриклеточные инфекты
(вирусы, внутриклеточные бактерии и простейшие) возникает КИО.
Доза антигена, кратность и путь его поступления, играют решающую
роль в возникновении иммунного ответа. Большие дозы антигена могут
индуцировать толерантность.
Антигенпрезентирующие клетки (АПК), процессируя и представляя
антиген вместе с молекулами HLA класса II Т-хелперам, обеспечивают
активацию Т-хелперов. В этом процессе важным и определяющим
является взаимодействие мембранных молекул Т-хелпера и АПК (СD40L
― СD40; CD 28 ― CD80 и CD86 молекул адгезии). Т-хелперы играют
регулирующую роль в выборе гуморального или клеточного иммунного
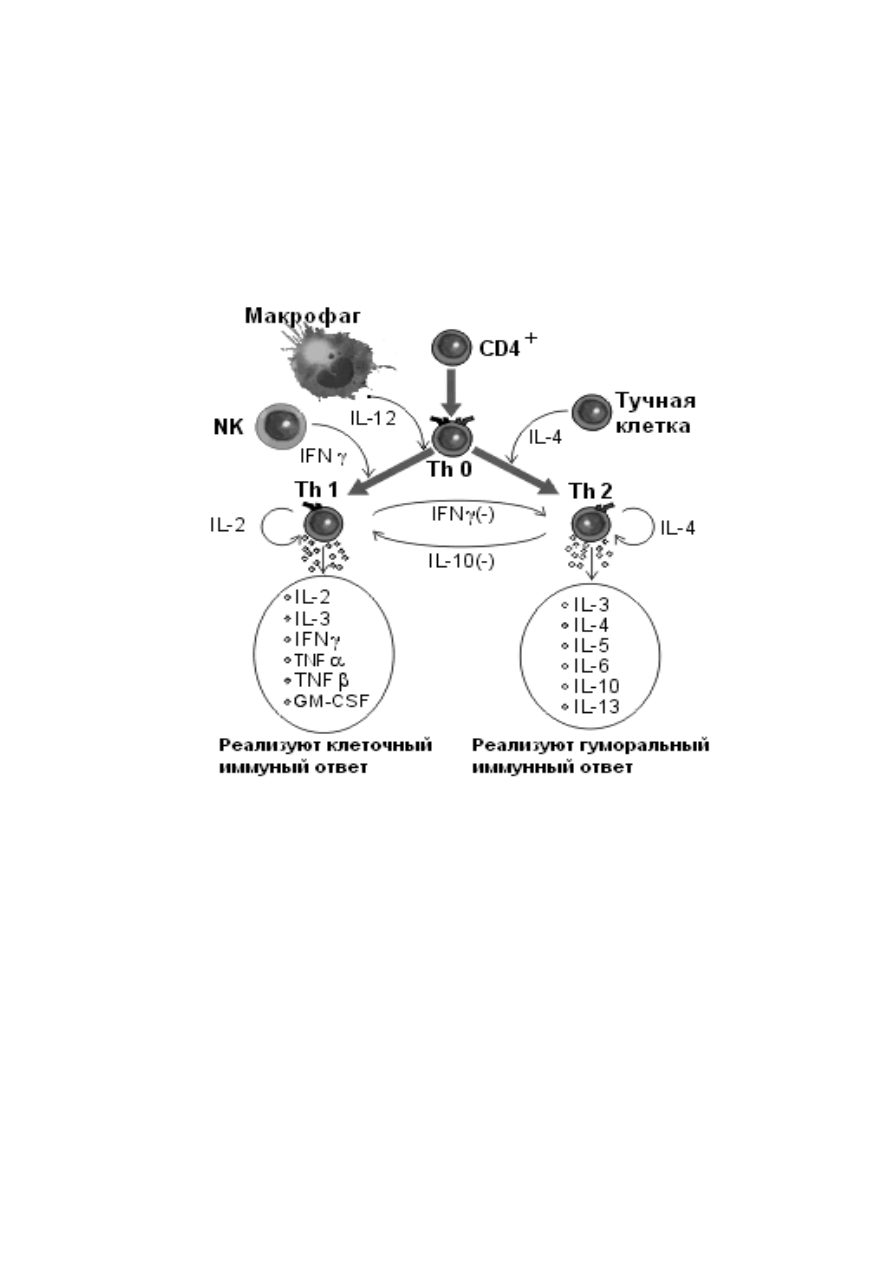
ответа (рисунок 8). Th1, продуцируя интерферон γ, подавляют развитие
Th2-клеток. И, наоборот, Th2, продуцируя ИЛ-4, ИЛ-10, подавляют Th1.
Таким образом, в процессе иммунного ответа преобладает клеточный или
гуморальный ответ.
Супрессия иммунного ответа наступает под действием множества
факторов. В первую очередь супрессия обусловлена элиминацией
антигена, исчезновением антигенной стимуляции и развитием апоптоза
простимулированных лимфоцитов. Отсутствие антигена в результате его
иммунной элиминации приводит к прекращению активации лимфоцитов
через TСR и BСR. Новые не иммунные лимфоциты не активируются, а в
проактивированных лимфоцитах снижается эспрессия антиапоптозных
генов, которые защищают лимфоциты от апоптоза в период иммунопоэза.
На лимфоцитах усиливается экспрессия CD95, рецепторов для
глюкокортикоидов и ФНО, что так же приводит к апоптозу.
102
Генетический контроль иммунного ответа осуществляет ГКГС. В
области ГКГС локализованы так называемые Ir (Immune response) гены,
определяющие степень интенсивности иммунного ответа на тот или иной
антиген. Таким образом, сила иммунного ответа на любой антиген у
каждого человека индивидуальна и детерминирована генетически.
Рисунок 8 ― кооперации ИКК в регуляции ИО
Активация В-лимфоцитов, отличия плазмоцита от В-лимфоцита.
Особенности иммунного ответа на Т-независимые антигены. Общая
схема гуморального иммунного ответа. Механизм эффекторного
действия антител и их характеристика
В процессе формирования В-клеток выделяют антигеннезависимую и
антигензависимую стадии.
Антигеннезависимая
стадия
созревания
В-лимфоцитов
не
определяется контактом с Аг, а зависит от микроокружения пре-В-клеток
(появляются под влиянием интерлейкинов ИЛ-3, ИЛ-4, ИЛ-6). На этой
стадии происходит формирование отдельных пулов генов, кодирующих
синтез Ig, и экспрессия этих генов. Однако на цитолемме пре-В-клеток еще
103
нет поверхностных рецепторов ― Ig (первичных Ig), компоненты
последних находятся в цитоплазме. Значительная часть пре-В-клеток
погибает в результате негативной селекции и апоптоза.
Образование В-лимфоцитов из пре-В-лимфоцитов сопровождается
появлением
на
их
поверхности
первичных Ig, способных
взаимодействовать с Аг. Только на этом этапе В-лимфоциты попадают в
кровоток
и
заселяют
периферические
лимфоидные
органы.
Сформировавшиеся молодые В-клетки накапливаются в основном в
селезенке, а более зрелые ― в лимфатических узлах.
Антигензависимая стадия созревания В-лимфоцитов начинается с
момента контакта этих клеток с Аг (в том числе ― аллергеном). В
результате происходит активация В-лимфоцитов, протекающая в два
этапа: пролиферации и диффе-ренцировки.
Пролиферация В-лимфоцитов обеспечивает два важных процесса:
• увеличение числа клеток, дифференцирующихся в продуцирующие
AT (Ig) В-клетки (плазматические клетки);
• образование В-лимфоцитов иммунологической памяти.
Процесс
антигензависимой
дифференцировки
В-лимфоцитов
находится под контролем интерлейкинов и протекает так:
• В-лимфоцит с помощью иммуноглобулиновых рецепторов
распознает и связывает антиген. Это стимулирует его переход из
покоящегося состояния (G0) в фазу G;
• в фазу G В-клетка увеличивается, на ее мембране образуются новые
рецепторы ― к фактору роста и к фактору дифференцировки;
• синтезируемые и секретируемые активированным Т-хелпером
факторы роста и дифференцировки взаимодействуют с рецепторами
В-клетки и переводят ее в следующую стадию клеточного цикла (S-
фазу) и в фазу дифференцировки;
• активированная
В-клетка
претерпевает 8-10 делений
и
одновременно дифференцируется в антителообразующую клетку ―
плазмоцит.
По мере созревания В-клеток и их превращения в плазматические
клетки происходит интенсивное развитие белоксинтезирующего аппарата,
комплекса Гольджи и исчезновение поверхностных первичных Ig. Вместо
них продуцируются уже секретируемые (т.е. выделяемые в биологические
жидкости ― плазму крови, лимфу, СМЖ и др.) антигенспецифические AT.
90-96% всего производимого плазмоцитом белка представляет антитело с
одинаковым активным центром против антигена, который вызвал
антигензависимую дифференцировку данного В-лимфоцита. Так возникает
104
клон антителообразующих клеток. Каждая плазматическая клетка
способна секретировать большое количество Ig ― несколько тысяч
молекул в секунду.
В-клетки
памяти
представляют
собой
долгоживущие
рециркулирующие малые лимфоциты. Они не превращаются в
плазматические клетки, но сохраняют иммунную «память» об Аг. Клетки
памяти активируются при повторной их стимуляции тем же самым Аг. В
этом случае В-лимфоциты памяти (при обязательном участии Т-хелперов и
ряда других факторов) обеспечивают быстрый синтез большого
количества специфических AT и развитие эффективного иммунного ответа
или аллергической реакции.
Гуморальный
иммунный
ответ
(ГИО)
―
это
сложная,
многокомпонентная
реакция
иммунной
системы
организма,
индуцированная антигеном (В-клеточными эпитопами) и реализуемая В-
системой лимфоцитов в кооперации с макрофагами и Т-хелперами. Это
взаимодействие находится под контролем HLA-системы. Кооперация
клеток реализуется через клеточные системы, поверхности, клеточные
рецепторы и клеточные медиаторы.
Тимуснезависимых антигенов мало. Они являются сильными
митогенами и имеют большое количество одинаковых эпитопов. На
поверхности В-лимфоцитов очень большое число антигенраспознающих
рецепторов одной специфичности. Как только на них действует
липополисахарид, происходит агрегация рецепторов (образование
«шапочки») ― это первый сигнал к активации В-лимфоцитов. Второй
сигнал В-лимфоциты получают от макрофага в виде ИЛ1. После этого
происходит активация В-лимфоцита и дифференциация их в
плазматические
клетки,
синтезирующие
иммуноглобулин
малой
специфичности IgМ.
Особенностью ИО в данном случае заключается в следующем:
• не
происходит
переключения
синтеза IgМ
на
синтез
иммуноглобулинов класса G и др. классов;
• тормозится ИО, т.к. не образуются клетки памяти;
• быстро возникает иммунологическая толерантность.
Распознание тимусзависимых антигенов идет в комплексе с
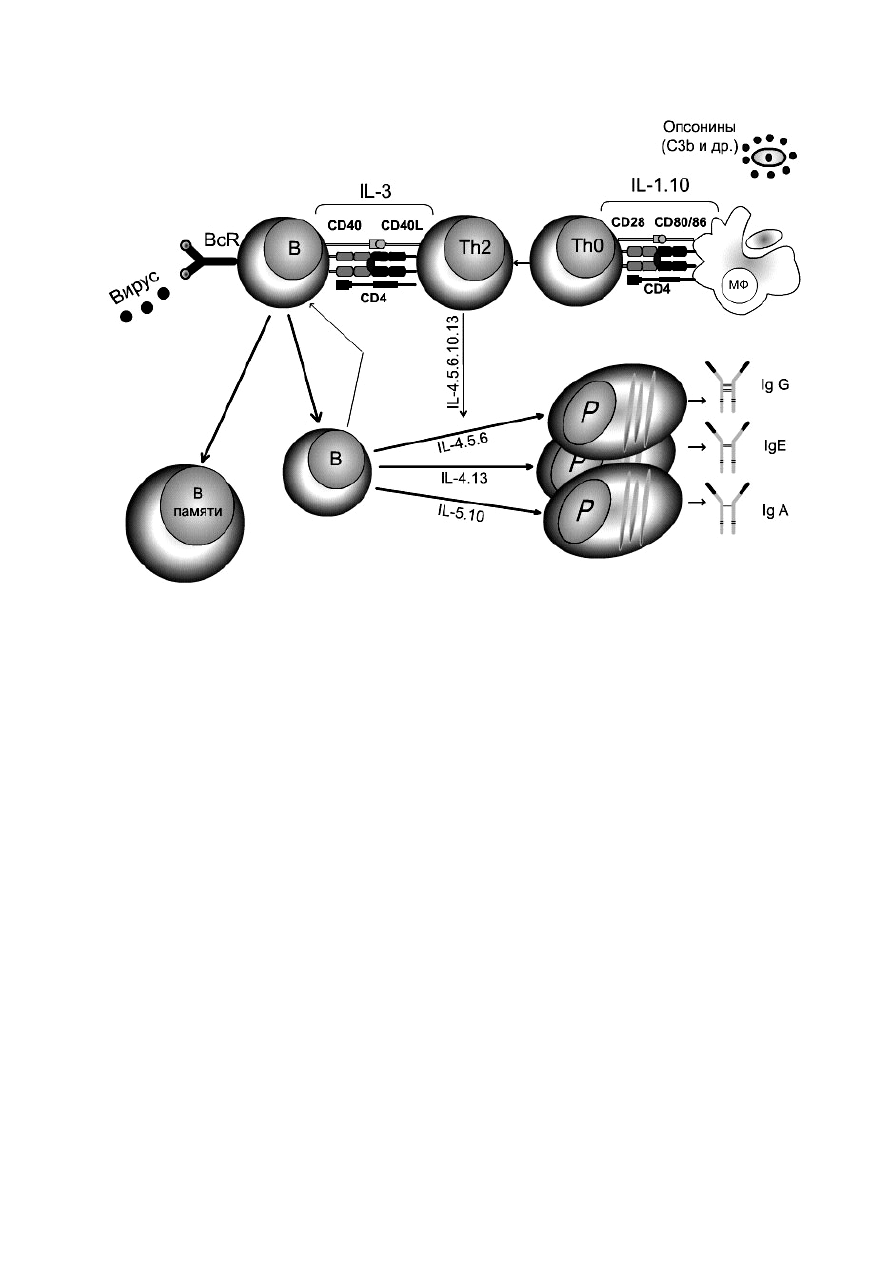
молекулами МНС. Схема ГИО приведена на рисунке 9.
105
Рисунок 9 ― Схема ГИО
Этапы ГИО:
1) поглощение антигена. Макрофаг движется в сторону АГ, реагируя на
положительные раздражители (С3а фрагмент активированного по
альтернативному
пути
комплемента),
т.е.
здесь
наблюдается
положительный хемотаксис ― «захват»;
2) процессинг антигена системой гидролитических ферментов
макрофагов до низкомолекулярных фрагментов;
3) представление антигена. Образовавшиеся комплексы белок МНС II
+ антигенный пептид (включает 12-25 аминокислотных остатков)
экспрессируются на мембране и представляются Т-хелперам (1-й сигнал).
Одновременно макрофаг активируется и выделяет фактор (ИЛ-1), который
также стимулирует Т-хелперы (2-й сигнал);
4) индуктивная фаза. Т-хелперы, получив два сигнала от макрофагов,
выделяют ИЛ-2, который стимулирует пролиферацию Т-лимфоцитов и В-
лимфоцитов. Причем активируются В-лимфоциты, имеющие мономерный
IgM в качестве рецептора, который соответствует этому антигену, т.е.
наступает селекция и избирательная стимуляция В-лимфоцитов;