ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.11.2019
Просмотров: 5764
Скачиваний: 159

УДК 58
ББК 28.57
Т22
Ответственный редактор член-
корреспондент РАН
А.И. Гречкин
Рецензенты:
доктор биологических наук, профессор
Л.Х. Гордон
доктор биологических наук, профессор
Л.П. Хохлова
ОТ АВТОРА
Тарчевский И.А.
Сигнальные системы клеток растений / И.А. Тарчевский; [Отв. ред. А.Н. Гречкин]. -
М.: Наука, 2002. - 294 с: ил. ISBN 5-02-006411-4
Рассматриваются звенья информационных цепей взаимодействия патогенов и растений, включающие
элиситоры, рецепторы элиситоров, G-белки, протеинкиназы и проте-инфосфатазы, факторы регуляции
транскрипции, репрограммирование экспрессии генов и ответ клеток. Главное внимание уделяется
анализу особенностей функционирования отдельных сигнальных систем клеток растений -
аденилатциклазной, МАР-киназной, фосфатидатной, кальциевой, липоксигеназной, НАДФН-оксидазной,
NO-
синтазной и протонной, их взаимодействию и объединению в единую сигнальную сеть. Предлагается
классификация патогениндуцированных белков по их функциональным признакам. Приводятся данные о
трансгенных растениях с повышенной устойчивостью к патогенам.
Для специалистов в области физиологии растений, биохимиков, биофизиков, генетиков,
фитопатологов, экологов, агробиологов.
По сети АК
Tarchevsky I.A.
Plant Cell Signaling Systems /1.A. Tarchevsky; [Ed. A.N. Grechkin]. - M.: Nauka, 2002. - 294
p.; il. ISBN 5-02-006411-4
The book discussed the members of signaling chains of interplay of pathogens and plant-host, namely elicitors,
receptors, G-proteins, protein kinases and protein phosphatases, transcription factors reprogramming of genes expression,
cell response. The main part of the book is devoted to functioning of separate cell signaling systems: adenylate cyclase,
MAP kinase, phosphatidate, calcium, lipoxy-genase, NADPH-oxidase, NO-synthase, protons systems. The concept of
interconnections of cell signaling systems and their integration to general cell signaling network is developing. The author
has preposed the classification of pathogen-related proteins according to their function properties. The data on transgenic
plants with the increased resistance to pathogens are presented.
For physiologists, biochemists, biophysicists, genetics, phytopathologists, ecologists, and agrobiologists
ISBN 5-02-006411-4
© Российская академия наук, 2002 ©
Издательство "Наука"
(художественное оформление), 2002
В последние годы стремительно развиваются исследования
молекулярных механизмов регуляции экспрессии генов под
влиянием изменения условий существования. В клетках растений
было обнаружено существование сигнальных цепей, которые с
помощью специальных белков-рецепторов, в большинстве случаев
расположенных в плазмалемме, воспринимают сигнальные
импульсы, преобразуют, усиливают и передают их в геном клетки,
вызывая репрограммирование экспрессии генов и изменения в
обмене веществ (в том числе кардинальные), связанные с
включением ранее "молчавших" и выключением некоторых
активных генов. Значимость сигнальных систем клеток была
продемонстрирована при изучении механизмов действия
фитогормонов. Была также показана определяющая роль
сигнальных систем в формировании адаптационного синдрома
(стресса), вызванного действием на растения абиотических и
биотических стрессоров.
Отсутствие обзорных работ, в которых анализировались бы
все звенья различных сигнальных систем, начиная с
характеристики воспринимаемых сигналов и их рецепторов,
преобразования сигнальных импульсов и передачи их в ядро и
кончая драматическими изменениями в обмене веществ клеток и их
структуре, заставили автора предпринять попытку восполнить этот
пробел с помощью предлагаемой вниманию читателей книги.
Необходимо учитывать, что исследование информационного поля
клеток еще очень далеко от завершения и многие детали его
структуры
и
функционирования
остаются
недостаточно
освещенными. Все это привлекает новых исследователей, для
которых обобщение публикаций по сигнальным системам клеток
растений будет особенно полезным. К сожалению, не все обзоры

статьи экспериментального характера вошли в список литературы, что в
определенной степени зависело от ограниченности объема книги и
времени для ее подготовки. Автор приносит извинения коллегам, чьи
исследования не были отражены в книге.
Автор выражает благодарость своим сотрудникам, принимавшим
участие в совместном исследовании сигнальных систем клеток
растений. Особую признательность автор выражает профессору Ф.Г.
Каримовой, кандидатам биологических наук В.Г. Яковлевой и Е.В.
Асафовой, А.Р. Муха-метшину и доценту Т.М. Николаевой за помощь в
подготовке рукописи к печати.
Работа выполнена при финансовой поддержке фонда Ведущей
научной школы РФ (гранты 96-15-97940 и 00-15-97904) и Российского
фонда фундаментальных исследований (грант 01-04-48-785).
ВВЕДЕНИЕ
Одной из важнейших проблем современной биологии является
расшифровка механизмов реагирования прокари-отических и
эукариотических организмов на изменения условий их
существования, особенно на действие экстремальных факторов
(стресс-факторов, или стрессоров), вызывающих у клеток
состояние стресса.
В процессе эволюции у клеток выработались приспособления,
позволяющие воспринимать, преобразовывать и усиливать
приходящие из окружающей среды сигналы химической и
физической природы и с помощью генетического аппарата
реагировать на них, не только адаптируясь к изменившимся
условиям, перестраивая свои обмен веществ и структуру, но и
выделяя различные летучие и нелетучие соединения во
внеклеточное пространство. Одни из них выполняют роль
защитных веществ против патогенов, другие могут рассматриваться
в качестве сигнальных молекул, вызывающих ответ других клеток,
расположенных на большом расстоянии от места действия на
растения первичного сигнала.
Можно считать, что все эти адаптивные события происходят в
результате изменений в информационном поле клеток. Первичные
сигналы с помощью различных сигнальных систем вызывают
реакцию со стороны генома клеток, проявляющуюся в
репрограммировании экспрессии генов. По сути дела, сигнальные
системы регулируют работу основного вместилища информации -
молекул ДНК. С другой стороны, они сами находятся под
контролем генома.
Впервые в нашей стране целенаправленно исследовать
сигнальные системы клеток начали Е.С. Северин [Северин,
Кочеткова, 1991] на животных объектах и О.Н. Кулаева [Кулаева
и др., 1989; Kulaeva,1990; Kulaeva et al., 1992; Кулаева, 1995;
Бурханова и др., 1999] - на растительных.

В представляемой вниманию читателей монографии содержится
обобщение результатов изучения влияния биотических стрессоров
на функционирование сигнальных систем клеток растений. В
настоящее
время
интенсивно
исследуются
МАР-киназная,
аденилатциклазная, фос-фатидатная, кальциевая, липоксигеназная,
НАДФН-окси-дазная, NO-синтазная и протонная сигнальные системы
и их роль в онтогенетическом развитии растений и в формировании
ответа на изменяющиеся условия существования, особенно на
действие различных абиотических и биотических стрессоров. Автор
решил сосредоточить внимание лишь на последнем аспекте этой
проблемы - на молекулярных механизмах ответа растений на
действие патогенов, тем более что в этот ответ вовлечен целый ряд
фито-гормонов и выяснение особенностей взаимодействия с ними
сигнальных систем клеток растений привлекает большое внимание
исследователей.
К патогенам (в широком понимании этого термина) относят не
только вирусы, бактерии и грибы, но и паразитирующие на растениях
нематоды и насекомые. Биотический стресс вызывают насекомые и
травоядные животные из-за появления у растений обширной раневой
поверхности, что не только само по себе приводит к стрессу, но и к
быстрому инфицированию раны патогенными вирусами, бактериями и
грибами.
Воздействие биотических стрессоров приводит к ответу растений, в
основных чертах сходному с ответом на абиотические стрессоры
[Neumann et al.,
1989; Тарчевский, 1993]. Он характеризуется
совокупностью неспецифических реакций, что и позволило называть
его адаптационным синдромом, или стрессом. Естественно, что могут
обнаруживаться и специфические черты ответа, зависящие от вида
стрессора, однако с усилением меры его воздействия на первый план
все в большей степени начинают выступать неспецифические
изменения [Меерсон, 1986; Тарчевский, 1993]. Наибольшее внимание
им было уделено Н.С. Введенским (представления о парабиозе), Д.С.
Насоновым и В.Я. Александровым (представления о паранекрозе), Г.
Селье [1972; и др.] - в работах, посвященных стрессу у
животных, В.Я. Александровым [1985] - в исследованиях молекуляр-
ных основ стресса.
К числу наиболее значительных неспецифических изменений
при биотическом стрессе можно отнести следующие:
1.
Фазность в развертывании во времени ответа на действие
патогена.
2.
Усиление катаболизма липидов и биополимеров.
3.
Повышение в тканях содержания свободных радикалов.
4.
Подкисление цитозоля с последующей активацией протонных
помп, что возвращает рН к исходному значению.
5.
Повышение в цитозоле содержания ионов кальция с
последующей активацией кальциевых АТФаз.
6.
Выход из клеток ионов калия и хлора.
7.
Падение мембранного потенциала (на плазмалемме).
8.
Снижение общей интенсивности синтеза биополимеров и
липидов.
9.
Прекращение синтеза некоторых белков.
10.
Усиление синтеза или синтез отсутствовавших так
называемых патогениндуцируемых защитных белков (хи-
тиназ, (3-1,3-глюканаз, ингибиторов протеиназ и др.).
11.
Интенсификация синтеза укрепляющих клеточные
стенки компонентов - лигнина, суберина, кутина, каллозы,
богатого оксипролином белка.
12.
Синтез антипатогенных нелетучих соединений -
фитоалексинов.
13.
Синтез и выделение летучих бактерицидных и фун-
гицидных соединений (гексеналей, ноненалей, терпенов и
Др->-
14.
Усиление синтеза и повышение содержания (или по
явление) стрессовых фитогормонов - абсцизовой, жасмо-
новой, салициловой кислот, этилена, гормона пептидной
природы системина.
15.
Торможение фотосинтеза.
16.
Перераспределение углерода из
|4
СО
2
, усвоенного в
процессе фотосинтеза, среди различных соединений -
уменьшение включения метки в высокополимерные соединения
(белки, крахмал) и сахарозу и усиление (чаще относи
тельное - в процентах от усвоенного углерода) - в аланин,
малат, аспартат [Тарчевский, 1964].
17.
Усиление дыхания с последующим его торможением.
Активация альтернативной оксид азы, изменяющей направленность
электронного транспорта в митохондриях.

18.
Нарушения ультраструктуры - изменение тонкой
гранулярной структуры ядра, уменьшение числа полисом и
диктиосом, набухание митохондрий и хлоропластов, умень
шение в хлоропластах числа тилакоидов, перестройка цито-
скелета [Neumann et al., 1989].
19.
Апоптоз (программируемая смерть) клеток, подверг
шихся воздействию патогенов, и соседних с ними.
20.
Появление так называемой системной неспецифиче
ской устойчивости к патогенам в удаленных от места
воздействия патогенов участках (например, метамерных
органах) растения.
Многие из перечисленных выше изменений являются следствием
"включения"
стрессорами
относительно
небольшого
числа
неспецифических сигнальных систем.
По мере все более глубокого изучения механизмов ответных
реакций растений на действие патогенов обнаруживаются новые
неспецифичные ответные реакции клеток растений. К ним относятся
и неизвестные ранее сигнальные пути.
При выяснении особенностей функционирования сигнальных
систем необходимо иметь в виду, что эти вопросы являются частью
более общей проблемы регуляции функционирования генома. Следует
заметить, что универсальность структуры основных носителей
информации клеток различных организмов - ДНК и генов -
предопределяет унификацию и тех механизмов, которые обслуживают
реализацию этой информации [Гречкин, Тарчевский, 2000]. Это
касается репликации ДНК и транскрипции, структуры и механизма
действия рибосом, а также механизмов регуляции экспрессии генов
изменяющимися условиями существования клеток с помощью набора в
значительной степени универсальных сигнальных систем. Звенья
сигнальных систем также в основном унифицированы (природа, найдя
в свое время оптимальное структурное и функциональное решение
биохимической или информационной задачи, сохраняет и тиражирует
его в процессе эволюции). В большинстве случаев самые
разнообразные химические сигналы, поступающие из окружающей
среды, улавливаются клеткой с помощью специальных "антенн" -
рецепторных белковых молекул, пронизывающих клеточную
мембрану и выступающих над ее поверхностями с наружной и внутрен-
ней стороны. Несколько типов строения этих рецепторов
унифицированы у клеток растений и животных. Некова-лентное
взаимодействие внешнего участка рецептора с той или иной
сигнальной молекулой, поступающей из среды, окружающей
клетку, приводит к изменению конформации рецепторного белка,
которое передается на внутренний, ци-топлазматический участок. В
большинстве
сигнальных
систем
с
ним
контактируют
посреднические G-белки - еще одно унифицированное (по своим
структуре и функциям) звено сигнальных систем. G-белки
выполняют функции преобразователя сигналов, передавая
сигнальный конформаци-онный импульс на стартовый фермент,
специфичный для той или иной сигнальной системы. Стартовые
ферменты одного типа сигнальной системы у различных
объектов также универсальны и имеют протяженные участки с
одной и той же последовательностью аминокислот. Одним из важ-
нейших унифицированных звеньев сигнальных систем являются
протеинкиназы (ферменты, переносящие концевой остаток
ортофосфорной кислоты с АТФ на те или иные белки),
активируемые продуктами стартовых сигнальных реакций или их
производными. Фосфорилированные с помощью протеинкиназ
белки являются следующими звеньями сигнальных цепей. Еще
одно унифицированное звено сигнальных систем клеток - это
белковые
факторы
регуляции
транскрипции,
которые
представляют собой один из субстратов протеинкиназных
реакций. Структура этих белков также в значительной степени
унифицирована,
а
модификации
структуры
определяют
принадлежность факторов регуляции транскрипции к той или иной
сигнальной системе. Фосфорилирование факторов регуляции
транскрипции обусловливает изменение конформации этих белков,
их активацию и последующее взаимодействие с промоторным
участком определенного гена, что приводит к изменению
интенсивности его экспрессии (индукции или репрессии), а в
крайних случаях - к "включению" некоторых молчавших генов
или "выключению" активных. Репрограммирование экспрессии
совокупности генов генома вызывает изменение соотношения
белков в клетке, что и является основой ее функционального
ответа. В отдельных случаях химический сигнал из внешней среды
может взаимодействовать с рецептором, расположенным внутри
клетки - в цитозоле или да-
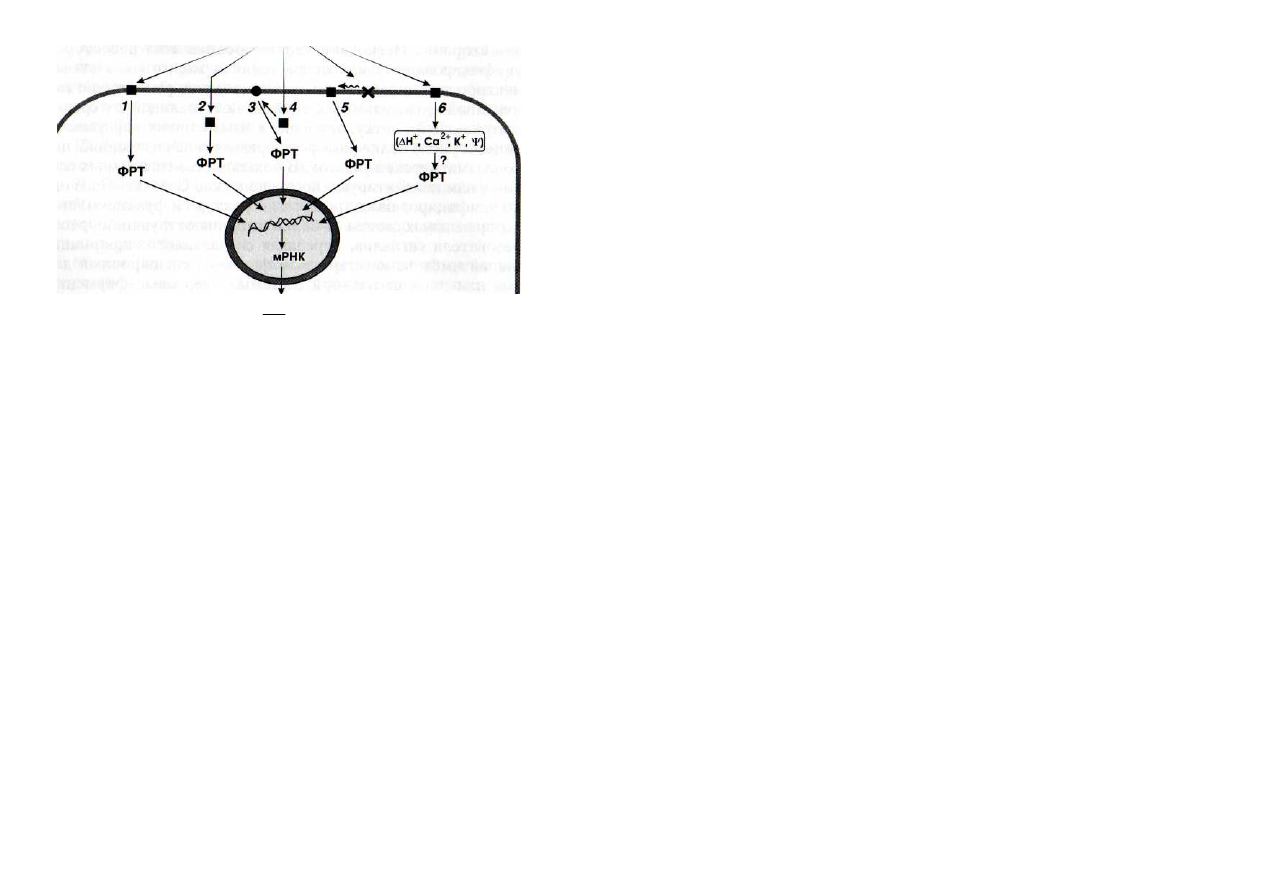
СИБ
Рис. 1. Схема взаимодействия внешних сигналов с рецепторами клетки
1 , 5 , 6 -
рецепторы, расположенные в плазмалемме;
2 , 4 -
рецепторы, находящиеся
в цитозоле;
3 -
стартовый фермент сигнальной системы, локализованный в
плазмалемме;
5 -
рецептор, активирующийся под влиянием неспецифического
изменения
структуры
липидной
составляющей
плазмалеммы;
СИБ
-
сигналиндуцированные белки; ФРТ -белковые факторы регуляции транскрипции; i|/ -
изменение мембранного потенциала
же ядре (рис. 1). В клетках животных такими сигналами являются,
например, стероидные гормоны. Этот информационный путь имеет
меньшее число интермедиатов, в связи с чем у него и меньше
возможностей для регуляции со стороны клетки.
В нашей стране всегда уделялось большое внимание проблемам
фитоиммунитета. Этой проблеме посвящен ряд монографий и обзоров
отечественных ученых [Сухоруков, 1952; Вердеревский, 1959; Вавилов,
1964; Горленко, 1968; Рубин и др., 1975; Метлицкий, 1976; Токин, 1980;
Метлиц-кий и др., 1984; Метлицкий, Озерецковская, 1985; Курсано-ва,
1988; Ильинская и др., 1991; Озерецковская и др., 1993; Кораблева,
Платонова, 1995; Чернов и др., 1996; Тарчев-ский, Чернов, 2000].
В последние годы особое внимание уделяется молекулярным
механизмам фитоиммунитета. Было показано, что
при инфицировании растений включаются различные сигнальные
системы, которые воспринимают, умножают и передают сигналы
от патогенов в генетический аппарат клеток, где происходит
экспрессия защитных генов, позволяющая растениям организовать
как структурную, так и химическую защиту от патогенов. Успехи
в этой области связаны с клонированием генов, расшифровкой их
первичной структуры (в том числе промоторных участков),
структуры кодируемых ими белков, использованием активаторов и
ингибиторов отдельных звеньев сигнальных систем, а также
мутантов и трансгенных растений с внедренными генами,
отвечающими за синтез участников рецепции, передачи и
усиления сигналов. В исследовании сигнальных систем клеток
растений важную роль играет конструирование трансгенных
растений с промоторами генов белков-участников сигнальных
систем.
В настоящее время сигнальные системы клеток растений при
биотическом стрессе наиболее интенсивно изучаются в Институте
биохимии им. А.Н. Баха РАН, Казанском институте биохимии и
биофизики РАН, Институте физиологии растений РАН,
Пущинском филиале Института биоорганической химии РАН,
центре "Биоинженерия" РАН, Московском и Санкт-Петербургском
государственных
университетах,
Всероссийском
научно-
исследовательском
институте
сельскохозяйственной
биотехнологии
РАСХН,
Всероссийском
научно-
исследовательском институте фитопатологии РАСХН и др.
Проблема расшифровки молекулярных механизмов био-
тического стресса, в том числе роли в его развитии сигнальных
систем, объединила на протяжении последних десяти с лишним лет
физиологов и биохимиков растений, микробиологов, генетиков,
молекулярных биологов, фитопатологов. Публикуется большое
количество экспериментальных и обзорных статей по различным
аспектам этой проблемы (в том числе в специальных журналах:
"Physiological and Molecular Plant Pathology", "Molecular Plant -
Microbe Interactions", "Annual Review of Plant Physiology and
Pathology").
В то же время в отечественной литературе отсутствует
обобщение работ, посвященных сигнальным системам клеток, что и
привело автора к необходимости написания предлагаемой
читателям монографии.
СИГНАЛЫ