ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.11.2019
Просмотров: 4028
Скачиваний: 98
Трансгенные растения с генами ферментов, нарушаю-
щих питание патогенов, насекомых и нематод.
Еще одно на-
правление работ по повышению устойчивости использует
свойство растений образовывать (в результате включения
сигнальных систем клеток) белки-ингибиторы протеиназ,
обеспечивающих азотное питание патогенных микроорга-
низмов, насекомых и нематод, или эндоглюканаз, участвую-
щих в их углеводном питании. С помощью переноса генов
белковых ингибиторов протеиназ [Johnson et al., 1989; De
Leo et al., 1998; Urwin et al., 1998; Walker et al.,
1999] и поли-
галактуроназ [Desiderio et al., 1997; Глинка, Проценко, 1998;
Devoto et al., 1998; Lorito et al.,
1998] достигалось подавление
развития этих организмов.
Трансгенные растения с генами хитиназ, Р-1,3-эндоглю-
каназ и лектинов.
Достаточно эффективное отражение ата-
ки патогенных грибов, насекомых и нематод осуществляет-
ся в результате привнесения в растения генов хитиназ из
других растений, насекомых или микроорганизмов [Neuhaus
et al., 1991; Hart et al., 1992; Ding et al., 1998; Datta et al., 2001;
Mora, Earle
2001], разрушающих хитин клеточных стенок
патогенных грибов или хитин насекомых. Опыты с транс-
генными растениями, в которые привносились в разной сте-
пени укороченные гены хитиназы, привели к выводу, что
in vivo
могут существовать различные формы хитиназ, об-
разующиеся благодаря протеолитическому процессингу
С-конца фермента и его гликозилированию [Zhu et al.,
2001]. Трансгенные растения с антисмысловым геном хити-
назы значительно уменьшали содержание этого защитного
белка [Samac, Shah, 1994]. В меньшей степени используются
гены Р-1,3-эндоглюканаз [Masoud et al., 1996; Borkowska
et al.,
1998], разрушающие полисахариды клеточных стенок
патогенных бактерий и грибов, или совместно гены этих
двух ферментов [Zhu et al., 1996]. Фунгицидной активностью
обладали растения табака с геном (3-1,3-1,4-глюканазы из
термофильной бактерии Clostridium thermocellum [Дарби-
нян и др., 1995]. Оказалось, что трансгенные растения с
привнесенным геном лектина обладают повышенной ус-
тойчивостью к повреждению личинками моли [Down et al.,
2001].
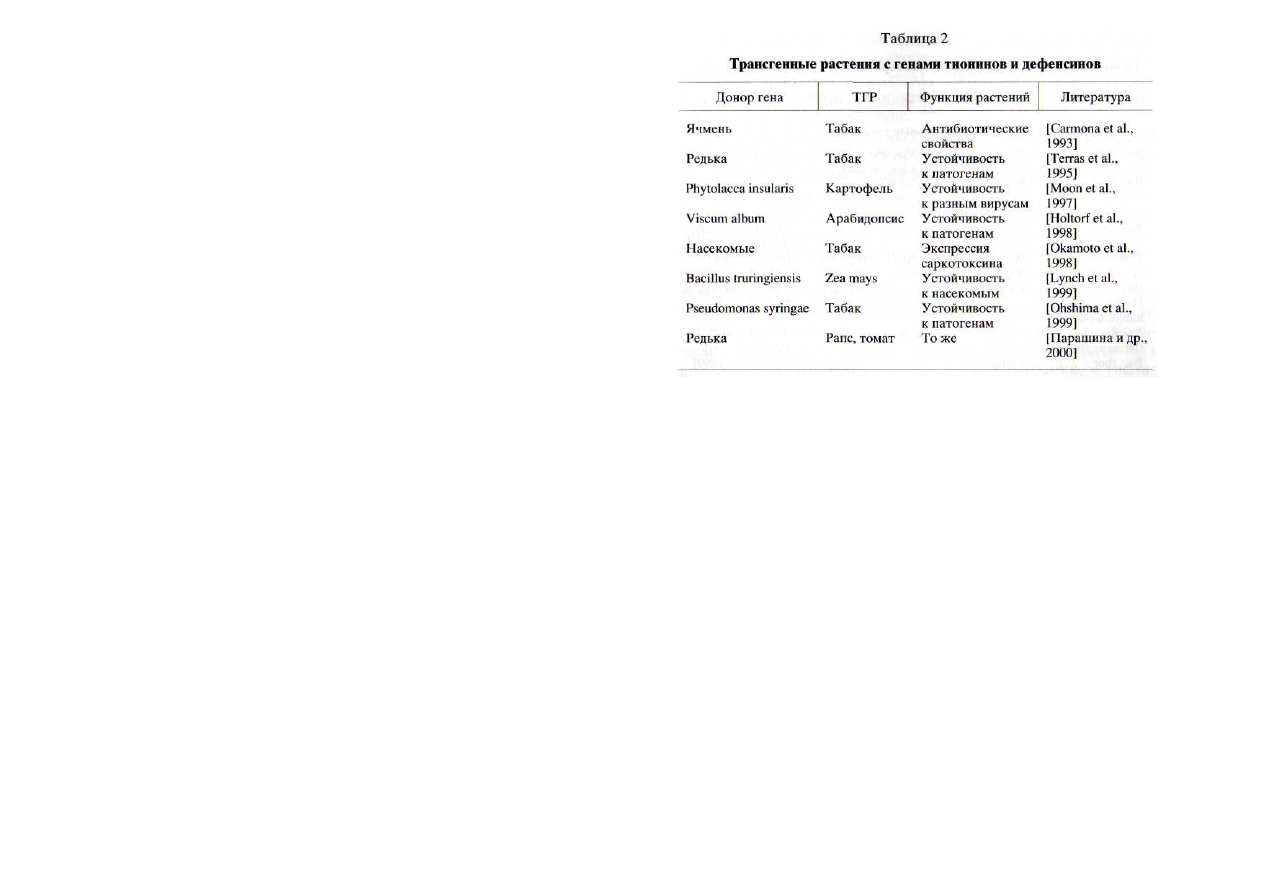
Трансгенные растения с генами дефенсинов, тионинов и
рибосомоинактивирующих белков.
Одним из важных на-
правлений создания устойчивых к патогенам растений явля-
ется перенос в них генов природных белков (дефенсинов,
тионинов, липидпереносящих белков), вызывающих нару-
шение функционирования клеточных мембран патогенных
грибов и бактерий [Carmona et al., 1993; Terras et al., 1995;
Butt et al., 1998; Dempsey et al., 1998; Gao et al.,
2000; Ляпко-
ва и др., 2001]. В роли поставщиков этих генов могут высту-
пать как сами растения, так и животные, насекомые, грибы
и бактерии (табл. 2), у которых, например, дефенсины име-
ют сходный план строения и достаточно консервативные
участки молекул [Zinn-Justin et al., 1996; Thevissen et al., 1997;
Kushmerick et al.,
1998]. В качестве примера можно привес-
ти трансгенные растения табака, обладающие повышенной
устойчивостью к патогенам, что было обеспечено перено-
сом гена дефенсина кролика [Zhang et al., 2000].
Еще одно направление создания устойчивых трансген-
ных растений - это перенос в них генов белков (N-гликози-
даз), нарушающих функционирование белоксинтезирую-
щей машины грибов и бактерий (так называемых рибосо-
моинактивирующих белков, блокирующих фактор элонга-
ции [Girbes et al., 1996; Hong et al., 1996; Lam et al., 1996;
Zoubenko et al., 2000; Nielsen, Boston,
2001]). Обнаружено,
90S
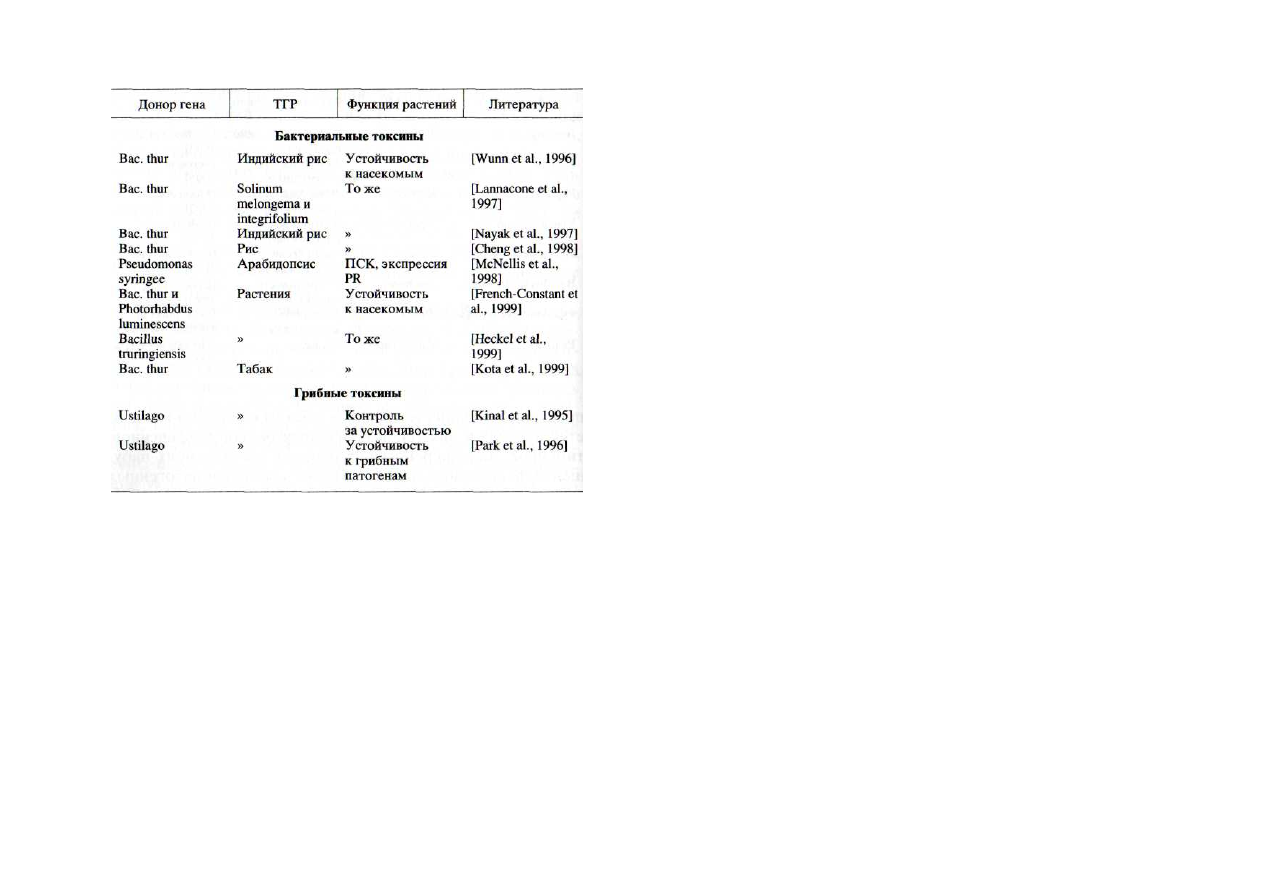
Таблица 3 Трансгенные растения с
генами бактериальных и грибных токсинов
что рибосомоинактивирующие белки действуют также на
вирусную РНК, что, по-видимому, связано с их РНКазной
активностью [Obrig et al., 1985; Mock et al., 1996].
Недавно были получены трансгенные растения с кон-
ститутивно экспрессируемыми привнесенными генами от-
носительно небольших белков пуроиндолинов, токсичных
для фитопатогенных грибов in vivo и in vitro [Krishnamurhy
etal., 2001].
К последним двум направлениям работ по созданию ус-
тойчивых трансгенных растений примыкает еще одно, в ко-
тором используются гены бактериальных токсинов, обла-
дающих инсектицидными свойствами (табл. 3).
Большой интерес вызывало установление факта индук-
ции синтеза РНК-зависимой РНК-полимеразы вирусами
или салициловой кислотой. Растения табака с привнесен-
ным геном антисмысловой РНК-зависимой РНК-полимера-
зы обнаруживали более высокое содержание вирусной
РНК и более ярко выраженные симптомы заболевания, чем
исходные растения [Xie et al., 2001]. Это означает, что
РНК-зависимая РНК-полимераза играет важную роль в ан-
тивирусной защите растений.
Из приведенных данных следует, что для создания
трансгенных растений, устойчивых к патогенным бактери-
ям, грибам и вирусам, а также к паразитирующим на расте-
ниях насекомым и нематодам используется целый арсенал
генов, кодирующих белки различных этапов функциониро-
вания сигнальных систем клеток растений, начиная с элиси-
торных белков и кончая антипатогенными, инсектицидны-
ми и антинематодными белками. Работы по созданию
трансгенных растений с повышенной устойчивостью к био-
тическим стрессорам осуществляются со все более высоки-
ми темпами и, безусловно, будут способствовать все боль-
шему расширению общей площади посевов, отведенных
для сельскохозяйственных трансгенных растений, и увели-
чению разнообразия видов и сортов этих растений [Lamb
et al.,
1992; Ноу, 1998; Salmeron, Vernooij, 1998; Бурьянов,
1999; Kolodziejczyk, Fedec,
1999; Романов, 2000; Borch,
Rasmussen, 2000; Melchers, Stuiver, 2000; Rommens, Kishore,
2000; Wolfenbarger, Phifer,
2000]. Опасность загрязнения не-
желательным генетическим материалом других хозяйствен-
но ценных растений (за счет переопыления, горизонтально-
го переноса генетического материала при инфицировании
растений вирусами и бактериями [Чернов и др., 1996] и т.д.),
связанная с повышением темпов создания трансгенных рас-
тений, заставляет исследователей учитывать эти факторы
при получении новых трансгенных сортов.

ЗАКЛЮЧЕНИЕ
Из приведенного в книге материала можно сделать вы-
вод о чрезвычайной сложности механизмов, формирующих
функциональный и структурный ответы клеток на внешние
сигналы. Под влиянием патогенов и элиситоров включают-
ся различные сигнальные системы растений и приходит в
движение вся сигнальная сеть клеток (напрашивается ана-
логия с паутиной, у которой импульс механического раздра-
жения передается к центру паутины не только по радиаль-
ным нитям - аналогам отдельных сигнальных систем, но и
по связывающим их поперечным нитям - аналогам сигналь-
ных интермедиатов). В результате осуществляются пере-
программирование работы генетического аппарата клеток
и формирование защитных антипатогенных химических и
физических барьеров.
Уже в процессе функционирования липоксигеназной,
НАДФН-оксидазной и NO-синтазной сигнальных систем
образуются очень активные антипатогенные соединения -
оксилипины (в том числе гексенали и ноненали), перекись
водорода, монооксид азота, пероксинитрит, способные по-
давлять развитие патогенов. Обращает на себя внимание,
что все эти соединения являются интермедиатами "кисло-
родных" сигнальных систем, у которых стартовые или
близкие к ним реакции осуществляются с участием кисло-
рода воздуха. Исключением является интермедиат адени-
латциклазной сигнальной системы - цАМФ, подавляющий
развитие патогенных грибов. Выше уже отмечалось, что
три перечисленные сигнальные системы могут быть отне-
сены к эволюционно наиболее "молодым". Перечисленные
выше интермедиаты эволюционно более "молодых" сиг-
нальных систем могут оказывать губительное действие не
только на клетки патогенов, но и на клетки растения-хозяина,
приводя к их апоптозу, что препятствует распростране-
нию патогенов из мест инфицирования в другие части рас-
тения.
Антипатогенным действием обладают также различные
элиситориндуцируемые "классические" фитоалексины -
фенилпропаноидные, летучие терпеноидные и другие со-
единения, образование которых начинается или интенсифи-
цируется за счет элиситориндуцируемого синтеза соответ-
ствующих ферментов. Повышение устойчивости к различ-
ным стрессорам, в том числе биотическим, определяется
также укреплением клеточных стенок (их лигнификацией,
синтезом каллозы и гидроксипролиновых белков, образо-
ванием сшивок между белками), изменением содержания и
соотношения белковых и липидных компонентов мембран.
Изменяется и ультраструктура клеток.
Исследование молекулярных механизмов взаимоотно-
шений патогенов и растений привело к появлению ряда пер-
спективных практических направлений формирования и
повышения фитоиммунитета. К ним относятся использова-
ние элиситоров и интермедиатов сигнальных систем в каче-
стве индукторов иммунитета и конструирование трансген-
ных устойчивых к патогенам растений с переносом в пос-
ледние генов белковых элиситоров, интермедиатов сиг-
нальных систем или защитных белков непосредственного
антипатогенного действия.
Дальнейшая расшифровка механизмов взаимодействия
сигнальных систем, по-видимому, должна составлять одну из
важнейших задач биохимии и клеточной биологии начала
XXI
в. Можно прогнозировать следующие направления ис-
следований и разработок, связанных с сигнальными система-
ми и сетями: поиск новых сигнальных систем и минорных
участников уже известных сигнальных систем; расшифровка
особенностей функционирования сигнальной сети как едино-
го целого; "привязка" отдельных сигнальных систем к тому
или иному виду сигналов и рецепторов; продолжение установ-
ления структуры промоторных участков и молекулярного
механизма их взаимодействия со "своими" факторами регу-
ляции транскрипции у различных генов; расшифровка меха-
низмов, обусловливающих временной (преходящий) харак-
тер включения тех или иных сигнальных систем и в то же
время длительную память об их включении, проявляющую-
ся, например, в формировании системного иммунитета против
патогенов; характеристика видовой, органной и тканевой
специфичности функционирования сигнальных систем; соз-
дание трансгенных форм растений с видоизмененными сиг-
нальными системами, что позволит получить или сверхчув-
ствительные формы нежелательных организмов (и поста-
вить их на грань выживания), или сверхустойчивые к биоген-
ным и абиогенным стрессорам; конструирование трансген-
ных форм растений с генами защитных белков, в том числе
прямого антипатогенного действия; использование приемов
генетической инженерии растений для получения ценных
фармакологических препаратов.
Необходимо отметить, что общие принципы работы
сигнальных систем в значительной степени универсальны.
Универсальность ДНК - основного вместилища информа-
ции, определяет сходство механизмов ее обслуживания в
клетках микроорганизмов, растений и животных. Это каса-
ется универсальности структуры рецепторов, встроенных в
клеточные мембраны, ассоциирующих с ними G-белков,
структуры стартовых ферментов сигнальных систем и фер-
ментов, ответственных за синтез и деградацию небелковых
вторичных посредников, структуры протеинкиназ, проте-
инфосфатаз, факторов регуляции транскрипции, РНК-по-
лимераз, рибосом и обслуживающих их работу белков.
Исследование особенностей функционирования сиг-
нальных систем клеток позволяет сформулировать следую-
щие общие положения [Гречкин, Тарчевский, 2000]: клетка
является многомерным информационным пространством,
образованным совокупностью взаимосвязанных сигналь-
ных систем и генома; существует постоянный двусторонний
обмен "командами" между геномом и сигнальными систе-
мами; оперативное управление жизнедеятельностью клетки
находится под контролем сигнальных систем. Сам по себе
геном является лишь хранилищем информации, реализуемой
с помощью сигнальных систем в зависимости от изменения
внутренней и окружающей клетку среды.
* * *
5-
7 июня 2001 г. в Москве был проведен Международ-
ный симпозиум по сигнальным системам клеток растений.
В решении симпозиума говорится, что в связи с бурным раз-
витием проблемы сигнальных систем клеток растений и с
первостепенной ее значимостью не только для фундамен-
тальной науки, но и для практических приложений в расте-
ниеводстве, биотехнологии и фармакологии необходимо
провести следующий симпозиум уже через три года.
Автор надеется, что публикуемая книга привлечет вни-
мание представителей различных научных направлений к
проблеме информационного поля клеток, и это приведет к
более эффективному ее решению.
СОДЕРЖАНИЕ
От автора
5
Введение
7
Патогены и элиситоры 14
Рецепторы элиситоров 27
G-
белки
35
Протеинкиназы и протеинфосфатазы
40
Факторы регуляции транскрипции
45
Промоторы генов белков сигнальных систем и защитных
белков 49
Аденилатциклазная сигнальная система 53
МАР-киназная сигнальная система
62
Фосфатидатная сигнальная система
67
Кальциевая сигнальная система 72
Липоксигеназная сигнальная система
84
НАДФН-оксидазная сигнальная система 103
NO-
синтазная сигнальная система
114
Протонная сигнальная система
123
Механизмы патогениндуцируемои смерти клеток 127
Сигнальная функция цитоскелета
134
Взаимодействие сигнальных систем со стрессовыми фито-
гормонами
139
Регуляция ионных потоков интермедиатами сигнальных
систем 149
Взаимодействие сигнальных систем
153
Патогениндуцируемые белки
170
Использование элиситоров и интермедиатов сигнальных
систем клеток для создания препаратов, повышающих устойчивость
растений к патогенам 190
Трансгенные растения с измененной устойчивостью к пато
генам 194
Заключение
208
Литература
212

Научное издание
Тарчевский Игорь Анатольевич
СИГНАЛЬНЫЕ СИСТЕМЫ КЛЕТОК РАСТЕНИЙ
Утверждено к печати
Ученым советом
Казанского института биохимии и биофизики
Казанского научного центра
Российской академии наук
Зав. редакцией
Н.А. Степанова
Редактор
Т.И. Белова
Художник
Е.А. Быкова
Художественный редактор
В.Ю. Яковлев
Технический редактор
О.В. Аредова
Корректоры
Г.В. Дубовицкая, А.В.
Морозова
ЛР N° 020297 от 23.06.1997
Подписано к печати 19.04.2002 Формат 60 х 9O'/i6. Гарнитура Тайме
Печать офсетная
Усл.печ.л. 18,5. Усл.кр.-отт. 19,0. Уч.-изд.л. 16,1 Тираж 810 экз. Тип. зак. 3275
Издательство "Наука"
117997 ГСП-7, Москва В-485, Профсоюзная ул., 90 E-mail: secret@naukaran.ru Internet:
www.naukaran.ru
Санкт-Петербургская типография "Наука" 199034, Санкт-Петербург В-34, 9-я линия, 12
И. А. Тарчевский
СИГНАЛЬНЫЕ
СИСТЕМЫ
КЛЕТОК
РАСТЕНИЙ
МОСКВА "НАУКА" 2002
За цикл работ по сигнальным системам клеток растений автор удостоен
премии им. А. Н. Баха Российской академии наук за 2002 г.