ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.11.2019
Просмотров: 5777
Скачиваний: 159

ПАТОГЕНЫ И ЭЛИСИТОРЫ
Болезни растений вызывают тысячи видов микроорганизмов,
которые можно разделить на три группы [Jackson, Taylor, 1996]:
вирусы (более 40 семейств) и вироиды; бактерии (Agrobacterium,
Corynebacterium, Erwinia, Pseudomonas, Xanthomonas, Streptomyces)
и
микоплазмоподобные
микроорганизмы;
грибы
(низшие:
Plasmodiophoromycetes,
Chitridomycetes,
Oomycetes:
высшие:
Ascomycetes, Basidi-omycetes, Deuteromycetes).
К патогенам относят также более 20 родов нематод, чаще всего
поражающих корни, в меньшей степени — ткани листьев.
Фитопатогенные нематоды являются облигатны-ми паразитами,
питающимися содержимым цитоплазмы живых клеток растений и в
связи с этим вызывающими обширные некрозы тканей растений
[Williamson, Hussey,
1996]. Структурами, обеспечивающими питание
нематод, являются секреторные гланды и стилет, приспособленный для
проникновения через клеточную стенку. Секреторная жидкость гланд,
содержащая гидролитические ферменты, освобождается через стилет в
периплазматическое пространство клетки. Целостность плазмалеммы
на начальных этапах процесса питания нематоды не нарушается. Она из-
влекает питательные вещества из цитозоля через небольшие поры
плазмалеммы, образующиеся в месте ее контакта с отверстием стилета.
Среди продуктов секреции гланд нематод обнаружены белки,
углеводы, ферменты целлюла-зы и протеиназы. Образующиеся в
результате контакта растения и нематоды элиситоры вызывают в
значительной степени неспецифичную ответную защитную реакцию
клеток хозяина, которая возникает вследствие "включения"
сигнальных систем атакуемых клеток [Williamson, Hussey, 1996]. Так,
нематоды индуцировали в клетках растений син-
тез защитных ферментов: фенилаланин-аммиак-лиазы
[Brueske,
1980] и анионной пероксидазы [Zacheo et al.,
1993].
Одним из важных компонентов экологических систем
являются растительноядные насекомые (взрослые и личиночные
формы), которые относятся в основном к подклассу высших, или
крылатых (Pterygota) [Шванвич, 1949]. Бескрылые формы,
относящиеся к этому подклассу, появились в результате утраты
этих органов в процессе эволюции крылатых форм. Подкласс
насчитывает 20 отрядов насекомых, среди которых имеются
полифаги, не обладающие специфичностью по отношению к
растению, олигофаги и монофаги, у которых ярко выражена
специфичность взаимодействия патогена и растения-хозяина.
Одни насекомые питаются листьями (всей листовой пластинкой
или скелети-руя лист), другие - стеблями (в том числе выгрызая
стебель изнутри), завязями цветов, плодами, корнями. Тли и цикады
высасывают сок из проводящих сосудов с помощью хоботка или
стилета.
Несмотря на принимаемые меры борьбы с насекомыми,
продолжает оставаться злободневной проблема уменьшения
причиняемого ими вреда. В настоящее время свыше 12% урожая
сельскохозяйственных растений на планете теряется в
результате атаки на них патогенных микроорганизмов,
нематод и насекомых [Rommens, Kishore, 2000].
Повреждение клеток приводит к деградации их содержимого,
например высокополимерных соединений, и появлению
олигомерных
сигнальных
молекул.
Эти
"обломки
кораблекрушения" [Тарчевский, 1993] достигают соседних клеток
и вызывают в них защитную реакцию, включающую изменение
экспрессии генов и образования кодируемых ими защитных
белков.
Часто
механическое
повреждение
растений
сопровождается их инфицированием, так как открывается раневая
поверхность, через которую в растение проникают патогены.
Кроме того, в ротовых органах насекомых могут обитать
фитопатогенные микроорганизмы. Известно, например, что
переносчиками микоплазмен-ной инфекции являются цикады, у
которых взрослые формы и личинки питаются соком ситовидных
сосудов растений, прокалывая хоботком-стилетом покровы
листьев и
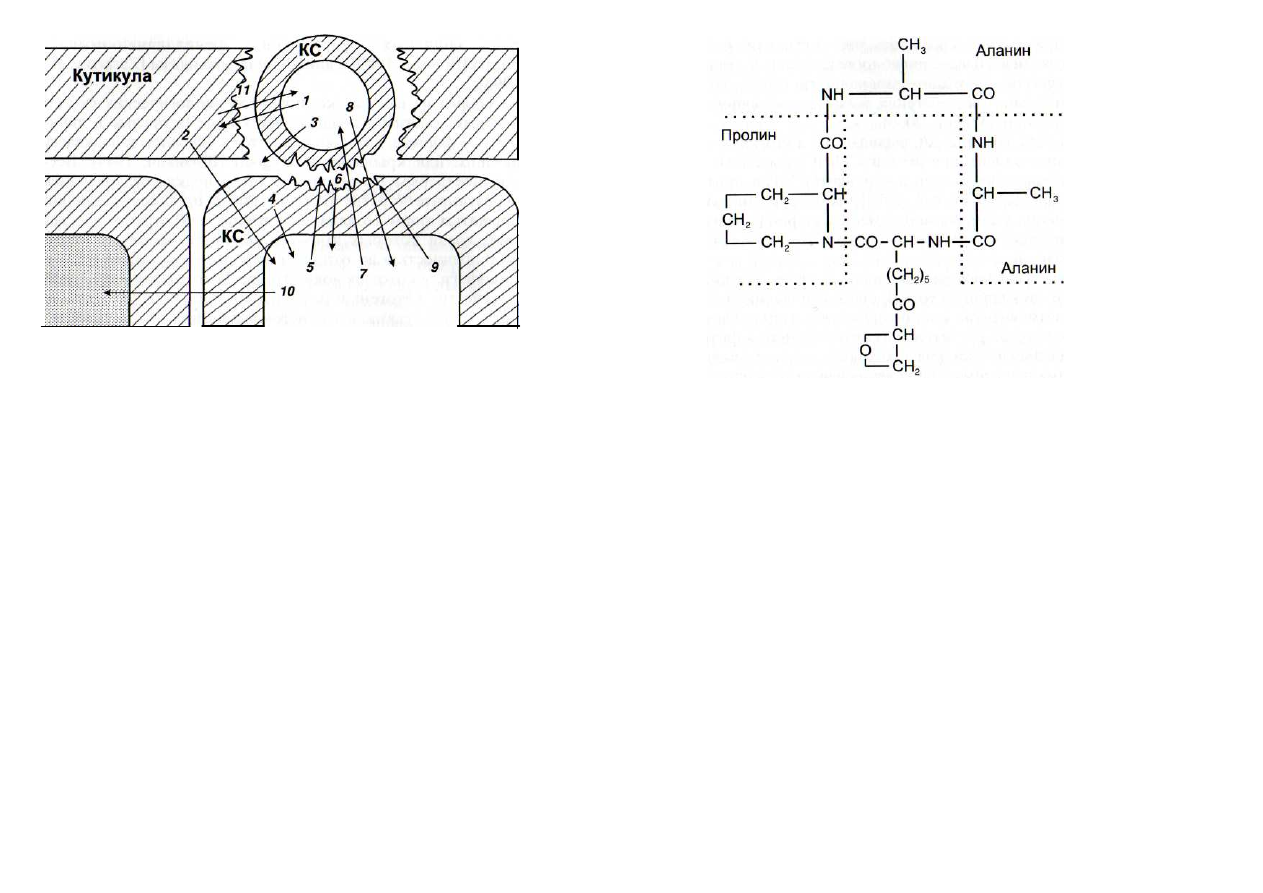
Рис. 2. Схема взаимодействия клетки патогена с растением-хозяином
/ -
кутиназа; 2 - продукты деградации компонентов кутикулы (возможно,
обладающие сигнальными свойствами);
3
- (3-
глюканаза и другие гликозилазы,
экскретируемые патогеном;
4
-
элиситоры - фрагменты клеточной стенки (КС)
хозяина;
5 -
хитиназы и другие гликозилазы, действующие разрушающе на КС
патогена;
6
-
элиситоры - фрагменты КС патогена; 7 - фитоалексины - ингибиторы
протеиназ, кутиназ, гликози-лаз и других ферментов патогена;
8
-
токсические
вещества патогена; 9 - укрепление КС хозяина за счет активации пероксидаз и усиления
синтеза лигнина, отложения оксипролиновых белков и лектинов;
10
-
индукторы
сверхчувствительности и некроза соседних клеток; // - продукты деградации кутина,
действующие на клетку патогена
молодых стеблей. Розанная цикадка, в отличие от других
представителей цикадовых, высасывает содержимое клеток. Цикады
производят меньшее повреждение тканей растений, чем листогрызущие
насекомые, тем не менее растения могут на него реагировать так же,
как на сопряженное с ним инфицирование растений.
При контакте с растениями клетки патогенов выделяют различные
соединения, обеспечивающие их проникновение в растение, питание и
развитие (рис. 2). Некоторые из этих соединений являются токсинами,
которые патогенные микроорганизмы выделяют для ослабления
сопротивляемости хозяина. В настоящее время описано более 20 хозяин-
специфичных токсинов, продуцируемых патогенными грибами.
Рис. 3. Фитотоксичное соединение из Cochlio-bolus
carbonum [Walton, 1996]
Бактерии и грибы образуют также неселективные токсины, в
частности фузикокцин, эрихосетен, коронатин, фазе-олотоксин,
сирингомицин, табтоксин [Walton, 1996].
Один из хозяин-специфичных токсинов, выделяемых
Pyrenophora triticirepentis, -
это белок 13,2 кДа, другие являются
продуктами вторичного метаболизма, имеющими самую
разнообразную структуру - это поликетиды, терпено-иды,
сахариды, циклические пептиды и т.д.
Как правило, к последним относятся пептиды, синтез которых
происходит вне рибосом и которые содержат остатки D-
аминокислот. Например, хозяин-специфичный токсин из
Cochliobolus
carbonum
имеет тетрапептидную циклическую
структуру
(D-npo-L-ana-D-ana-L-A3JJ),
где последняя аббревиатура
означает 2-амино-9,10-эпокси-8-оксо-де-каноевую кислоту [Walton,
1996; 2000] (рис. 3). Токсин образуется в клетках патогена с
помощью токсинсинтазы. Устойчивость к этому соединению у
кукурузы зависит от гена, кодирующего НАДФН-зависимую
карбонил-редуктазу, восстанавливающую карбонильную группу,
что приводит к

деактивации токсина. Оказалось, что в организме растения-хозяина
токсин вызывает ингибирование гистон-деацетилаз и, как следствие,
сверхацетилирование гистонов. Это подавляет защитный ответ
растения, вызываемый инфицированием патогенами [Walton, 2000].
Другой тип соединений, выделяемых патогенами, получил название
элиситоров (от англ. elicit - выявлять, вызывать). Собирательный
термин "элиситор" был предложен впервые в 1972 г. [Keen et al., 1972]
для обозначения химических сигналов, возникающих в местах
инфицирования растений патогенными микроорганизмами, и получил
широкое распространение.
Элиситоры играют роль первичных сигналов и приводят в действие
сложнейшую сеть процессов индукции и регуляции фитоиммунитета.
Это проявляется в синтезе защитных белков, нелетучих растительных
антибиотиков - фитоалек-синов, в выделении антипатогенных летучих
соединений и др. В настоящее время охарактеризована структура
множества природных элиситоров. Некоторые из них продуцируются
микроорганизмами, другие (вторичные элиситоры) образуются при
ферментативном расщеплении высокополимерных соединений
кутикулы и полисахаридов клеточных стенок растений и
микроорганизмов,
третьи
представляют
собой
стрессовые
фитогормоны, синтез которых в растениях индуцируется патогенами и
абиогенными стрессорами. К числу важнейших элиситоров относятся
белковые соединения, экскретируемые патогенными бактериями и
грибами, а также белки оболочки вирусов [Toedt et al., 1999]. Наиболее
изученными белковыми элиситорами можно считать небольшие (10
кДа), консервативные, гидрофильные, обогащенные цистеином
элиситины, секретируе-мые всеми исследовавшимися видами
Phytophthora
и Pythium [Kamoun et al., 1993; Pernollet et al., 1993;
Pallaghy et al., 1994; Huet et al., 1995; Lloyd, 1995; Panabieres et al., 1995;
Kamoun et al., 1998; Zhang et al.,
1998]. К ним относится, например,
криптогеин [Keller et al.,1999; Lebrun-Garcia et al., 1999; Foissner et al.,
2000].
Элиситины вызывают сверхчувствительность и отмирание
инфицированных клеток, особенно у растений рода Nicotiana [Kamoun
et al.,
1997]. Наиболее интенсивное образование фитофторой элиситинов
происходит при росте ми-
целия гриба и на последних стадиях инфекции, когда наблюдаются
обильная споруляция гриба и некрозы в листьях. Интересно, что у
паразитирующего на томатах гриба Cladosporium fulvum
пептидный элиситор не образовывался в оптимальных условиях in
vitro,
но интенсивно синтезировался внутри листа растения [Van
den Ackerveken et al., 1994].
Обнаружено, что элиситины способны переносить сте-ролы
через мембраны, так как имеют стеролсвязывающий сайт [Mikes et
al.,
1998]. Многие патогенные грибы сами не могут синтезировать
стеролы, что делает понятной роль элиситинов не только в питании
микроорганизмов, но и в индуцировании защитной реакции растений.
Из фитофторы был выделен гликопротеидный элиситор 42 кДа [Jabs
et al., 1997; Hirt, Scheel,
2000]. Его активность и связывание с
белковым рецептором плазмалеммы, мономерная форма которого
представляет собой белок 100 кДа [Nennstiel et al., 1998],
обеспечивалась олигопептидным фрагментом из 13 аминокислотных
остатков. Расоспецифичный элиситорный пептид, состоящий из 28
остатков аминокислот с тремя дисуль-фидными группами, удалось
получить из фитопатогенного гриба Cladosporium fulvum [De Wit,
1997; Van den Hooven et al.,
1999], причем образовывался пептид из
предшественника, содержавшего 63 аминокислоты. Этот фактор
авиру-лентности обнаруживал структурную гомологию с рядом
небольших пептидов, таких как ингибиторы карбоксипеп-тидазы и
блокаторы ионных каналов [Pallaghy et al., 1994], и связывался
рецепторным белком плазмалеммы, по-видимому, вызывая его
модуляцию, димеризацию и передачу сигнального импульса в
сигнальные системы [De Wit, 1997]. Из более крупного пре-протеина
Cladosporium fulvum,
состоящего из 135 аминокислот, в ходе
посттрансляционного процессинга образуется элиситорный белок,
насчитывающий
106
аминокислот.
Элиситорные
белки,
продуцируемые ржавчинным грибом Uromyces vignae, представляют
собой два небольших полипептида 5,6 и 5,8 кДа, по свойствам не-
похожие на другие элиситины [De Silva, Heath, 1997]. Среди
бактериальных белковых элиситоров наиболее изучены харпины
[Hahn, 1996; Desikan et al., 1998; Pontier et al., 1998; Xie, Chen, 2000].
Многие фитопатогенные бактерии продуцируют элиситорные
олигопептиды (созданы их синтетиче-

ские аналоги), соответствующие наиболее консервативным участкам
белка - флагеллина [Felix et al., 1999; Gomez-Gomez et al., 1999],
являющегося важным фактором вирулентности этих бактерий. Из
Erwinia amylovora
выделен новый элиситорный белок, С-область
которого гомологична ферменту пектатлиазе, способной вызывать
появление эли-ситорных олигомерных фрагментов - продуктов
деградации пектина [Gaudriault et al., 1998]. Патогенная бактерия
Erwinia carotovora
экскретирует элиситорный белок харпин и ферменты
пектатлиазу,
целлюлазу,
полигалактуроназу
и
протеазы,
гидролизующие полимерные компоненты клеточных стенок растения-
хозяина (см. рис. 2), в результате чего образуются олигомерные
элиситорные молекулы [Y. Liu et al., 1998]. Интересно, что
пектатлиаза, выделяемая Erwinia chrysanthemi [Shevchik et al., 1998],
приобретала активность в результате внеклеточного процессинга.
Некоторые липиды и их производные также относятся к
элиситорам, в частности 20-углеродные полиненасыщенные жирные
кислоты некоторых патогенов - арахидоно-вая и эйкозапентаеновая
[Ильинская и др., 1991; Озерец-ковская и др., 1993; Озерецковская,
1994; Гилязетдинов и др., 1995; Ильинская и др., 1996а, б; Ильинская,
Озерец-ковская, 1998], и их оксигенированные производные. В об-
зорной работе [Ильинская и др., 1991] обобщаются данные об
элиситорном действии на растения липидов (липопро-теинов),
продуцируемых патогенными грибами. Оказалось, что элиситорным
эффектом обладает не белковая часть липопротеинов, а их липидная
часть, представляющая собой не свойственные для высших растений
арахидоно-вую (эйкозатетраеновую) и эйкозопентаеновую кислоты.
Они вызывали образование фитоалексинов, некротиза-цию тканей и
системную устойчивость растений к различным патогенам. Продукты
липоксигеназного превращения в тканях растений С
20
жирных кислот
(гидроперокси-, гидрокси-, оксо-, циклические производные,
лейкотрие-ны), образующиеся в клетках растения-хозяина с помо-
щью ферментного липоксигеназного комплекса (субстратами
которого могут быть как С,
8
, так и С
20
полиеновые жирные кислоты),
оказывали сильнейшее влияние на защитную реакцию растений. Это
объясняется, по-видимому, тем, что в неинфицированных растениях
нет оксиге-
нированных производных 20-углеродных жирных кислот, и их
появление
в
результате
инфицирования
приводит
к
драматическим результатам, например к образованию некрозов
вокруг инфицированных клеток, что создает барьер для
распространения патогенов по растению.
Имеются данные, что индуцирование патогеном липо-
ксигеназной активности приводило к формированию ответной
реакции растения и в том случае, когда элиситор не содержал С
20
жирных кислот и субстратом липоксигеназной активности могли
быть только собственные С
18
полиеновые жирные кислоты, а
продуктами - октадеканоиды, а не эйкозаноиды. Элиситорными
свойствами обладают также сиринголиды [Л et al., 1998] и
цереброзиды - сфинголипид-ные соединения [Koga et al., 1998].
Цереброзиды А и С, изолированные из Magnaporthe grisea, были
наиболее активными элиситорами для растений риса. Продукты
деградации цереброзидов (метиловые эфиры жирных кислот,
сфинго-идные основания, гликозил-сфингоидные основания) не об-
наруживали элиситорной активности.
Некоторые элиситоры образуются в результате действия на
ткани растений гидролаз, выделяемых патогенами. Назначение
гидролаз двоякое. С одной стороны, они обеспечивают питание
патогенов, необходимое для их развития и размножения, с другой -
разрыхляют
механические
барьеры,
стоящие
на
пути
проникновения патогенов в места их обитания в растениях.
Одним из таких барьеров является кутикула, состоящая главным
образом из гетерополимера кутина, погруженного в воск.
Обнаружено более 20 мономеров, из которых состоит кутин
[Kolattukudy, Soliday, 1985; Airansinen, Paaso,
1990]. Это различной
длины насыщенные и ненасыщенные жирные кислоты и спирты, в
том
числе
гидроксилированные
и
эпокси-дированные,
дикарбоксиловые длинноцепочечные кислоты и т.д. В кутине
большинство первичных спиртовых групп участвует в образовании
эфирных связей, так же как часть вторичных спиртовых групп,
обеспечивающих сшивки между цепями и точки ветвления в
полимере. Часть другого "барьерного" полимера - суберина, по
составу близка к кутину. Главное его отличие в том, что свободные
жирные кислоты являются основным компонентом субериновых
восков, в то время как в кутине их очень мало. Кроме того, в
суберине

присутствуют главным образом С
22
и С
24
жирные спирты, в то время как в
кутине - С
26
и С
28
[Kolattukudy,
1987]. Для преодоления поверхностного
механического барьера растений многие патогенные грибы выделяют
ферменты, гидролизу-ющие кутин и часть составляющих суберина.
Продуктами кутиназной реакции были различные оксигенированные
жирные кислоты и спирты [Kolattukudy, 1985], в основном 10,16-
дигидрокси-Ск,- и 9,10,18-тригидрокси-С
|8
-
кислоты, представляющие
собой сигнальные молекулы, индуцирующие в прорастающей споре
гриба образование и выделение дополнительных количеств кутиназы,
"разъедающих" кутин и облегчающих проникновение гриба внутрь
растения. Было обнаружено, что лаг-период появления у гриба
кутиназной мРНК после начала образования упомянутых выше ди- и
триоксикислот составляет всего 15 мин, а выделения дополнительной
кутиназы - в два раза больший. Повреждение гена кутиназы у Fusarium
solani
сильно снижало степень вирулентности этого гриба [Kolattukudy et
al.,
1995]. Ингибирование кутиназы с помощью химических препаратов
или антител предотвращало инфицирование растений. Предположение о
том, что оксигенированные продукты деградации кути-на могут
выступать в роли не только индукторов образования кутиназы у
патогенов, но и элиситоров защитных реакций у растения-хозяина
[Тарчевский, 1993], впоследствии подтвердилось [Fauth et al., 1998].
После проникновения патогенных микроорганизмов через кутикулу
одни из них перемещаются в проводящие пучки растений и используют
для своего развития имеющиеся там питательные вещества, а другие
транспортируются внутрь живых клеток хозяина. В любом случае
патогены встречаются с еще одним механическим барьером - клеточ-
ными стенками, состоящими из различных полисахаридов и белков и в
большинстве случаев укрепленными жестким полимером - лигнином
[Тарчевский, Марченко, 1987; Tarchevsky, Marchenko, 1991]. Как уже
упоминалось выше, для преодоления этого барьера и обеспечения
своего развития углеводным и азотным питанием патогены выделяют
ферменты, гидролизующие полисахариды и белки клеточных стенок.
Специальные исследования показали, что при взаимодействии
бактерий и тканей растения-хозяина ферменты
деградации появляются не одновременно. Например, пек-
тилметилэстераза присутствовала и в неинокулированных
бактерией Erwinia carotovora subsp. atroseptia тканях клубней
картофеля [Pagel, Heitefuss, 1990], тогда как полигалактуро-
назная, пектатлиазная, целлюлазная, протеазная и ксила-назная
активности появлялись соответственно через 10, 14, 16, 19 и 22 ч
после инокуляции.
Оказалось, что олигосахаридные продукты деградации
полисахаридов клеточных стенок растений обладают эли-
ситорными свойствами. Но активные олигосахариды могут
образовываться и полисахаридами, входящими в состав
клеточных стенок патогенов. Известно, что одним из способов
защиты растений от патогенных микроорганизмов является
образование после инфицирования и выделение за пределы
плазмалеммы ферментов - хитиназы и β-1,3-глюканазы,
гидролизующих полисахариды хитин и β-1,3-полиглюканы
клеточных стенок патогенов, что приводит к подавлению их роста
и развития. Обнаружено, что олигосахаридные продукты такого
гидролиза являются и активными элиситорами защитных
реакций растений. В результате действия олигосахаридов
повышается устойчивость растений к бактериальной, грибной или
вирусной инфекции [Ryan, 1987; Albersheim et al.,1992; Doares et
al., 1995b; Bohland et al., 1997].
Олигосахаридным элиситорам, их строению, активности,
рецепторам, "включению" ими сигнальных систем клеток,
индукции экспрессии защитных генов, синтезу фитоалексинов,
реакции сверхчувствительности и другим ответам растений
посвящен целый ряд обзорных статей [Ryan, 1987; Albersheim et al.,
1992; Ebel,
1998; и др.].
В лаборатории Элберсгейма [Albersheim, 1969; Элберс-гейм,
Дарвилл, 1985; и др.], а затем в ряде других лабораторий показано,
что
олигогликозиды,
образующиеся
в
результате
патогениндуцированной
эндогликозидазной
деградации
гемицеллюлоз и пектиновых веществ растений, хитина и хитозана
грибов, могут играть роль биологически активных веществ. Было
даже предложено считать их новым классом гормонов
("олигосахаринов", в отличие от олигосахаридов, не обладающих
активностью). Образование олигосахаридов в результате гидролиза
полисахаридов, а не в ходе синтеза из моносахаридов было
показано на примере