ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.11.2019
Просмотров: 4025
Скачиваний: 98

ТРАНСГЕННЫЕ РАСТЕНИЯ
С ИЗМЕНЕННОЙ УСТОЙЧИВОСТЬЮ
К ПАТОГЕНАМ
Успехи молекулярной генетики, появление эффектив-
ных методов переноса в растения "чужих" существующих
в природе и искусственно конструируемых генов, клониро-
вание генов, изучение структурных и функциональных
особенностей генов и их промоторных участков обеспечи-
ли большие возможности в создании трансгенных расте-
ний специально для изучения особенностей функциониро-
вания сигнальных систем клеток растений. Кроме того,
трансгенные сельскохозяйственные растения с привнесен-
ными генами элиситоров, интермедиатов сигнальных сис-
тем и элиситориндуцируемых белков оказались более ус-
тойчивыми к патогенам и стали использоваться в практи-
ческих целях. В настоящее время ими заняты значитель-
ные посевные площади. Работы в этой области продвига-
ются настолько интенсивно, что появилась необходимость
систематизировать их и ознакомить с ними специалистов,
работающих в области физиологии растений и смежных
направлениях науки.
Эффективность работ по созданию трансгенных расте-
ний в значительной степени определялась результатами ис-
следований молекулярных основ адаптации и иммунитета,
соответственно, к абиотическим и особенно к биотическим
стрессорам.
При изучении особенностей влияния патогенных микро-
организмов на растения было обнаружено образование в
атакуемых тканях патогениндуцируемых защитных белков
(PR) [Neumann et al., 1989; Stintzi et al.,
1993]. Оказалось, что
эта реакция в определенной степени неспецифична: многие
защитные белки образуются при атаке на растения самых
разнообразных патогенных грибов, бактерий, вирусов, а
также насекомых и паразитирующих на растениях круглых
червях - нематодах. Последнее можно рассматривать и как
приспособительную реакцию против последующего инфи-
цирования растений через образовавшуюся раневую по-
верхность.
Патогены, продуцируемые ими и инфицированными
растениями элиситоры, "включают" сигнальные системы
клеток [Тарчевский, 2000], которые осуществляют рецеп-
цию, преобразование и усиление элиситорного сигнала и
передачу его в геном, где происходит экспрессия генов за-
щитных белков, что обеспечивает появление локальной и
системной устойчивости. В предыдущем разделе все пато-
ген(элиситор)индуцируемые белки были подразделены на
несколько групп по тем функциям, которые они выполня-
ют [Тарчевский, 2001].
Для создания устойчивых к биогенным стрессорам
трансгенных растений используются гены белков - участ-
ников сигнальных путей клеток. К ним относятся белковые
элиситоры; интермедиаты сигнальных систем, например
ферменты, катализирующие синтез или деградацию вто-
ричных мессенджеров; белки, обеспечивающие устойчи-
вость самого растения-хозяина или нарушение функций па-
тогеных микроорганизмов, растительноядных насекомых
или нематод.
Очевидно, что чем более раннее звено сигнальной цепи
клетки растения кодирует переносимый ген (например, ген
элиситорного белка), тем менее специфичным и более раз-
нообразным будет набор изменений в функционировании
трансгенного растения, обеспечивающих появление повы-
шенной устойчивости к стрессорам. Можно ожидать, что
более направленной и специфичной будет защита растений,
вызванная переносом в них генов белков прямого антипато-
генного действия, являющихся заключительным звеном
функционирования сигнальных цепей.
Обычно практикуется прикрепление переносимого ге-
на к промотору другого гена растения (рис. 55) - к консти-
тутивно функционирующему или, чаще всего, эффектив-
но изменяющему активность под влиянием воздействия на
растения того или иного стрессора. Иногда с целью боль-
шего повышения устойчивости трансгенного растения в

Рис. 55. Схема строения генов,
используемых в трансгенных
растениях
1 -
промоторная область;
2 —
соответствующий
промотору
"свой" ген;
3 -
чужеродный ген;
4
и
5 -
два чужеродных гена под
контролем одного промотора;
6 -
репортерный ген;
6
и 7 - два ре-
портерных гена под контролем
одного промотора; 2 и 7 - "свой" и
репортерный гены под контролем
одного промотора
него могут переносить не один, а различные гены [Zhu
et al., 1996].
Трансгенные растения с репортерными генами.
Для ис-
следования сигнальных систем используют трансгенные
растения, у которых к промотору гена, установленного ра-
нее или предполагаемого участника сигнальной системы,
прикрепляется так называемый репортерный ген, кодирую-
щий белок, активность которого может быть определена,
например белок, обладающий способностью люминесциро-
вать в присутствии определенных люминофоров. Чаще все-
го репортерный ген присоединяется к промоторному участ-
ку исследуемого гена. Интенсивность экспрессии репортер-
ных генов и распределение по органам растения или внутри
клеток кодируемых ими белков достаточно легко исследо-
вать по люминесценции. В большинстве случаев в качестве
репортерного используется ген фермента (3-глюкуронида-
зы (GUS) из энтеробактерии Е. coli. Фермент гидролизует
Р-£)-глюкурониды, превращая их в D-глюкуроновую кисло-
ту и агликоновый фрагмент, но может также отщеплять от
полисахаридов остаток (3-глюкуроновой кислоты, связан-
ный с сахарами [Gilissen et al., 1998]. Фермент проявляет
активность в форме гомотетрамера, обычно локализуется в
цитоплазме, но с помощью введения в репортерный ген по-
следовательности нуклеотидов, кодирующей транспортный
пептид, можно позволить ферменту перенос через мембра-
ны и накопление, например, в эндоплазматической сети.
Качественный, количественный и гистохимический анали-
зы активности GUS проводятся с помощью коммерческих
препаратов глюкуронидов по люминесценции продуктов
реакции в присутствии люминофоров. Этот подход оказал-
ся эффективным при изучении сигнального пути, "включа-
емого" элиситорными белками - элиситинами. Оказалось,
что молекула элиситина имеет два отличающихся участка
структуры, один из которых определяет сигнальный путь,
ведущий к синтезу защитных белков, а другой - к индукции
некрозов [Perez et al., 1997]. Репортерный ген GUS приме-
нялся также для определения структуры промоторов, на-
пример, отвечающих за экспрессию генов этилениндуциру-
емых защитных белков [Eyal et al., 1993], для изучения осо-
бенностей регуляции сигналиндуцируемого синтеза супер-
оксиддисмутазы [Herouart et al., 1994], каталазы [Guan,
Scandalios,
1993], анионных пероксидаз и их локализации в
различных органах растений [Mohan et al., 1993a, b; Klotz
et al., 1998; Gray-Mitsumune et al.,
1999], обогащенных гид-
роксипролином белков клеточных стенок [Wycoff et al., 1995;
Puigdomenech et al., 1997], msr (multiple stimulus response)
ге-
нов, от которых зависит апоптоз [Pontier et al., 1998], осмо-
тинов [Zhu et al., 1995; 1996], дефенсинов [Manners et al.,
1998; Mitter et al., 1998], PR1
защитных белков [Tornero et al.,
1997], протеаз [Jorda, Vera, 2000], хитиназ [Clarke et al., 1994;
Leah et al., 1994; Shinshi et al.,
1995], р-1,3-глюканаз
[Castresana et al., 1990; Vogeli-Lange et al., 1994; Alonso et al.,
1995], стриктозидин-синтазы - ключевого фермента элиси-
ториндуцируемого образования терпеноидных алкалоидов
[Memelink et al.,
1999], а также ферментов синтеза фенил-
пропаноидных фитоалексинов - фенилаланин-аммиак-лиа-
зы и сесквитерпенциклазы [Yin et al., 1997], 4-кумарат-ко-
фермент А-лигазы [Hauffe et al., 1991]. Введение гена (3-
глюкуронидазы помогло понять, как распределяется по
органам и тканям калретикулиновый ген [Coughlan et al.,
1997], изменяющий активность в ходе развития растений, и
моносахаридный Н
+
-
симпортерный ген [Truernit et al., 1996],
экспрессируемый при действии патогенов и элиситоров.
С помощью гена GUS удалось установить, что экспрессия
белка оболочки вируса, обеспечивающего его транспорти-
ровку по растению, локализована в тканях флоэмы вегета-
тивных органов [Christou et al., 2000]. Это объясняет, поче-
му скорость распространения вируса по растению значи-
тельно увеличивается после его попадания во флоэму из
клеток мезофилла листа [Курсанов, 1976]. Использование
гена (3-глюкуронидазы, введенного под промоторы двух
изоформ супероксиддисмутазы [Tanaka et al., 1995], показа-
ло, что только одна из них активируется экзогенной абсци-
зовой кислотой. При изучении особенностей экспрессии ге-
на тримерной протеинфосфатазы типа 2А, играющей важ-
ную роль в регуляции функционирования сигнальных сис-
тем, GUS-метод позволил выяснить, что от набора катали-
тического и регуляторных субъединиц протеинфосфатазы
зависит ее специфичность, активность и распределение внут-
ри клетки [Thakore et al., 1999].
Для исследования роли ионов кальция в функциониро-
вании сигнальных систем используют трансгенные расте-
ния с привнесенным конститутивно экспрессируемым ге-
ном экворина (aequorin) [Knight et al., 1991; Chandra et al.,
1997; и др.] - Са
2+
-
зависимого флуоресцентного белка (из
представителя кишечнополостных - Aequorea victoria), ко-
торый образуется в результате посттранскрипционной мо-
дификации из предшественника - апоэкворина 22 кДа. Эк-
ворин локализован в цитозоле клеток, и интенсивность его
люминесценции в присутствии люминофора коэлентерази-
на зависит от концентрации ионов кальция в этом компарт-
менте. Найдена также принципиальная возможность раз-
мещения экворина в таких органоидах, как митохондрии и
хлоропласты, с помощью конструирования химерных генов
апоэкворина с прикрепленными к ним фрагментами, коди-
рующими синтез сигнальных последовательностей, от ко-
торых зависит транспорт белков в органоиды. Были найде-
ны аналоги коэлентеразина, которые делают экворин
более чувствительным и позволяют определять низкие кон-
центрации ионов кальция [Knight et al., 1993].
Для исследования сигнальных систем клеток растений
и особенностей взаимодействия патогенов и растений ис-
пользуют еще один репортерный ген из Aequorea victoria -
ген так называемого зеленого флуоресцентного белка
[Rossi et al., 1996; Liu, Kolattukudy, 1999; Liu et al., 2001].
Практикуется получение гетерорепортерных генных кон-
струкций, кодирующих как этот белок, так и (3-глюкуро-
нидазу, под промотором одного и того же гена [Quaedvlieg
et al., 1998; Ottenschlager et al.,
1999], что предоставляет до-
полнительные возможности исследования распределения
и транспортировки белков, а также взаимоотношений па-
тогенов и растений. С помощью гена зеленого флуорес-
центного белка, введенного в геном растений табака, уда-
лось установить, что вирус мозаики табака транспортиру-
ется по растению с помощью двух различных механиз-
мов - медленного, когда вирус передвигается от клетки к
клетке, что сопровождается репликацией, и быстрого, осу-
ществляющегося по сосудам растения [Casper, Holt, 1996].
Использование зеленого флуоресцентного белка под про-
мотором белка движения вируса X картофеля позволило
получить новую информацию о движении вирусов через
плазмодесмы [Oparka et al., 1996].
В некоторых опытах определялась активность репор-
терных генов хлорамфеникол-ацилтрансферазы, например,
при исследовании особенностей действия патогенов и эли-
ситоров на активность халконсинтазы [Loake et al., 1991;
Yamada et al., 1994] -
ключевого фермента синтеза фенил-
пропаноидных фитоалексинов.
Использование репортерного гена люциферазы для ис-
ледования сигнальных систем клеток растений поставлено
под сомнение в связи с тем, что субстрат этого гена (люци-
ферин) сам вызывает активацию генов защитных белков
IJorda, Vera, 2000].
Трансгенные растения с репортерными генами доста-
точно часто конструируют для познания структуры промо-
горных участков генов белков - участников сигнальных си-
стем клеток растений, ферментов синтеза фитоалексинов и
защитных белков.
Трансгенные растения с генами белков-элиситоров.
На-
ибольшее число работ в этом направлении посвящено соз-
данию более устойчивых растений с помощью переноса в
них генов белков оболочки вирусов [Miller, Hemenway,
199
8], элиситирующих включение сигнальных систем кле-
ток путем взаимодействия с рецепторами (табл. 1) [Глагоц-
кая и др., 1990а, б; Culver, Dawson, 1991; Lomonossoff, 1995;
Bendahmane et al., 1997; Hammond, Dienelt, 1997; Lim et al.,
1997; Moon et al., 1997; Seppanen et al., 1997; Watanabe et al.,
1997; Chowrira et al., 1998; Guo et al., 1998; Lorito et al., 1998;
Miller, Hemenway, 1998; Moreno et al., 1998; Ravelonandro
et al., 1998; Reimann-Philipp, 1998; Beachy et al., 1999; Boria
et al., 1999; Han et al., 1999; Jan et al., 1999; Keller et al., 1999;
Kehm et al., 2001; Savenkov, Valkonen, 2001; Sunter et al.,
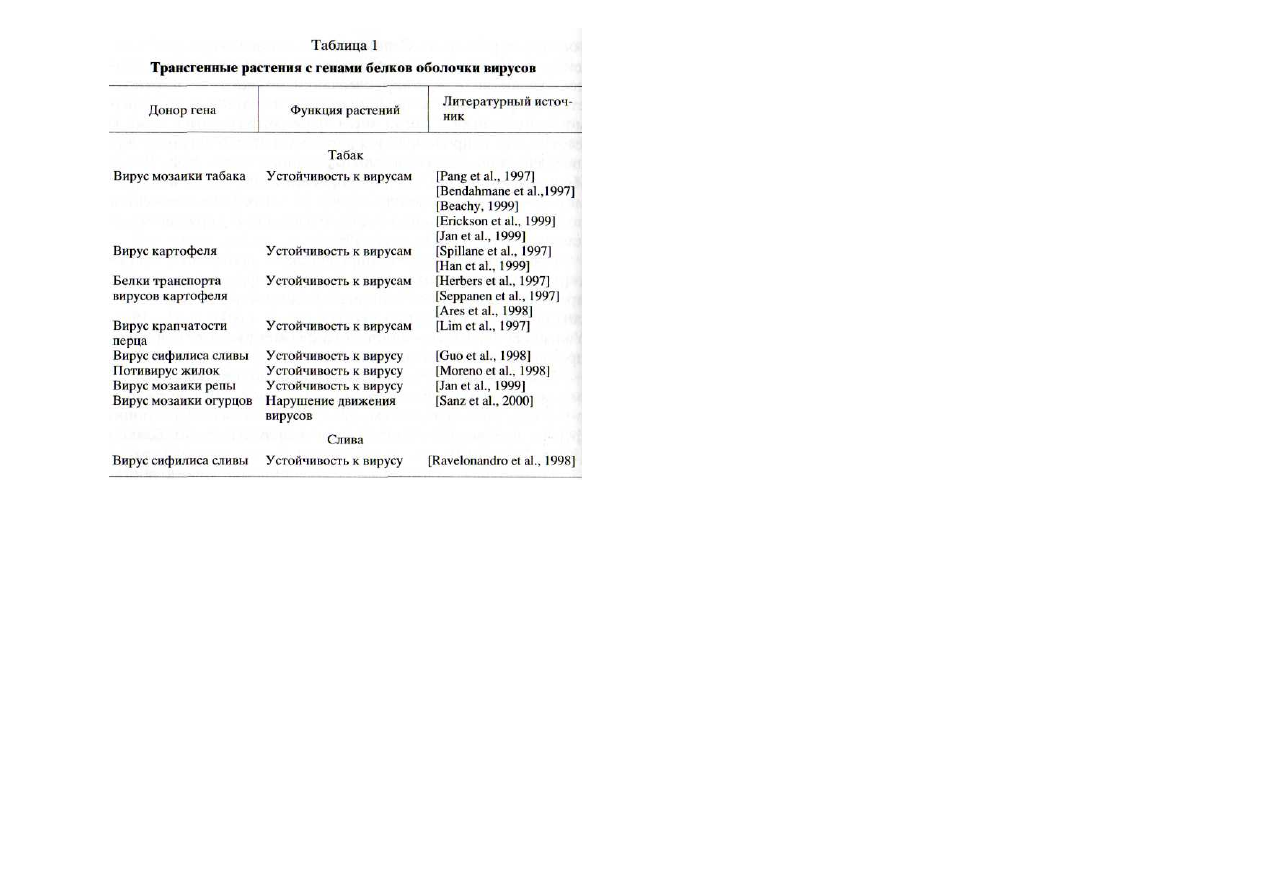
2001]; вирусных белков, обеспечивающих их транспорти-
ровку из клетки в клетку [Herbers et al., 1997; Seppanen et al.,
1997; Spillane et al., 1997; Sanz et al., 2000; Kotlizky et al., 2001;
Simone et al.,
2001] и, наконец, вирусных репликаз и геликаз
[Jones et al., 1998; Wittner et al., 1998; Erickson et al., 1999; Guo
et al., 1999; Thomas et al., 2000; Vazquez et al.,
2001], а также
рибозимов [Kwon et al., 1997; Yang et al., 1997].
Сконструировано относительно мало трансгенных рас-
тений с генами белков элиситоров, продуцируемых пато-
генными грибами и бактериями. Из них особенно большое
внимание исследователей привлекает криптогеин фито-
фторы [Lorito et al., 1998; Keller et al., 1999].
Трансгенные растения с генами белков, участвующих в
синтезе стрессовых фитогормонов.
Стрессовые фитогор-
моны (абсцизовая, жасмоновая и салициловая кислоты, эти-
лен, системин) могут рассматриваться в качестве соедине-
ний, элиситирующих сигнальные системы, поэтому пред-
ставляют интерес работы, в которых приводятся данные о
трансгенных растениях с привнесенными генами белков,
участвующих в образовании фитогормонов [Hedden,
Phillips,
2000]: просистемина и системина [McGurl et al., 1992,
1994а], 1-аминоциклопропан-1-карбоксилат-синтазы [С. Liu
et al., 1998; De Martinis, Mariani, 1999] -
одного из ферментов
синтеза этилена, алленоксидсинтазы [Harms et al., 1995;
С. Wang et al., 1999; Laudert et al., 2000], катализирующей об-
разование предшественника жасмоната.
Трансгенные растения с генами белков - участников
сигнальных систем. В
настоящее время осуществлен пере-
нос в растения генов белков, участвующих почти во всех
этапах передачи сигнала от элиситоров до генома клеток.
Получены трансгенные растения с рецепторным автоки-
назным белком [Ohtake et al., 2000; Tao et al., 2000] и с рядом
белков, принимающих участие в обслуживании сигнальных
систем клеток. Одним из важнейших участников этих сис-
тем являются G-белки и протеинкиназы. Трансгенные рас-
тения с активным геном G-белка подтверждают это поло-
жение, обнаруживая повышенную устойчивость к вирусам
[Sano et al., 1994; Sano, Ohashi, 1995; Li et al.,
2001], так же
как растения с привнесенным геном протеинкиназы
|Morello et al.,
2000]. Были получены трансгенные растения
с привнесенным геном кальцийзависимой протеинкиназы
[Romeis et al.,
2000]. Обязательным участником сигнальных
систем клеток являются факторы регуляции транскрипции,
и трансгенные растения с активными генами этих белков
обнаруживают повышенную устойчивость к патогенам
|Mayda et al., 1999; Park et al.,
2001]. Недавно было обнару-
жено, что при вирусной инфекции повышается активность
гена РНК-зависимой РНК-полимеразы растения табака,
что предположительно повышало устойчивость к вирусу.
Использование трансгенных растений табака с антисмыс-
ловым геном этого фермента показало, что у исследуемых
растений наблюдалось нарушение защитных реакций и как
локальное (в месте инфицирования), так и системное накоп-
ление вирусов [Xie et al., 2001].
Выше уже были охарактеризованы основные сигналь-
ные системы клеток растений: аденилатциклазная, МАР-
киназная, фосфатидатная, кальциевая, липоксигеназ-ная,
супероксиддисмутазная (НАДФН-оксидазная), NO-син-
тазная и протонная сигнальные системы.
Изменение работы МАР-киназной сигнальной системы
и повышение устойчивости к стрессорам достигалась в
трансгенных растениях с помощью гена МАРК [Seoa et al.,
1999] или изоформы МАРККК, активируемой перекисью
водорода [Kovtun et al., 2000], снижение интенсивности син-
теза фосфатидной кислоты в фосфатидатной системе у ара-
бидопсиса - с помощью антисмыслового подавления синте-
за фосфолипазы Д [С. Wang, 2000]. В кальциевой сигналь-
ной системе использовались трансгенные растения с анти-
смысловым геном фосфолипазы С [Sanchez, Chua, 2001],
привнесенными генами кальмодулина [Harding et al., 1997] и
инозитол-5'-фосфатазы (снижающей содержание инози-
толтрисфосфатов и инозитолтетракисфосфатов, что подав-
ляет кальциевый сигнальный путь) [Berdy et al., 2001;
Sanchez, Chua,
2001], в липоксигеназной - генами гидро-
пероксидлиазы [Vancanneyt et al., 2001], образующей анти-
патогенные летучие С
6
-
соединения, и алленоксидсинтазы
[Harms et al.,
1995], катализирующей образование фитодие-
новой кислоты, из которой синтезируется жасмоновая кис-
лота. Трансгенные растения арабидопсиса с привнесенным
геном метилтрансферазы обнаруживали повышение устой-
чивости к патогенным грибам за счет усиления образования
из жасмоновой кислоты подвижной стрессовой сигнальной
молекулы метилжасмоната [Seo et al., 2001], что вызвало
усиление формирования системного иммунитета.
Особенно много работ посвящено трансгенной модифи-
кации супероксидсинтазной сигнальной системы с помо-
щью переноса генов супероксиддисмутазы [Van Camp et al.,
1996; Chung et al., 2000; Pan et al.,
2001], катал азы [Takahashi
et al., 1997; Willekens et al., 1997; Chamnongpol et al., 1998;
Mittler et al.,
1999], аскорбатпероксидазы [Mittler et al., 1999]
или гидроксилазы салициловой кислоты [Chamnongpol et al.,
1998; Reuber et al., 1998; Kumar, Klessig, 2000; Lee et al., 2001;
Yoshioka et al.,
2001], катализирующей ее превращение в не-
активный катехол. Как правило, генетическое вмешатель-
ство в функционирование НАДФН-оксидазной системы
приводило к изменениям в вызванном патогенами апоптозе
клеток. Трансгенные растения табака с перенесенным в них
геном подавления апоптоза животных проявляли большую
устойчивость к вирусу табачной мозаики [Mitsuhara et al.,
1999; Dickman et al.,
2001], что еще раз свидетельствует об
общности механизмов апоптоза и его участников. Ингиби-
тор апоптоза животных клеток оказывал аналогичное дейст-
вие и на клетки арабидопсиса [Kawai-Yamada et al., 2001]. На
протекание апоптоза и устойчивость к патогенам может
оказывать влияние активность не только супероксидисму-
тазы и каталазы, но и ферментов, от которых также зави-
сит окислительно-восстановительный режим клеток — цис-
теин-пероксиредоксина и глутатион-пероксидазы, о чем
свидетельствуют трансгенные растения с генами этих фер-
ментов [Lee et al., 2000; Roxas et al., 2000]. Интересны
результаты опытов с катионной пероксидазой растений, по-
казавшие, что ее свойства можно изменять с помощью на-
правленной генетической замены одной из трех гликозили-
рующихся аминокислот [Lige et al., 2001].
В связи с тем, что Н
+
-
АТФаза, по-видимому, играет важ-
ную роль в функционировании протонной сигнальной сис-
темы [Schaller, (Decking, 1999], представляют интерес дан-
ные о повышении устойчивости растений к грибам и бакте-
риям после введения в них гена бактериородопсина из
Halobacterium halobium [Mittler et al., 1995; Abad et al., 1997;
Rizhsky, Mittler,
2001], выполняющего функции протонной
помпы, как было показано в серии работ В.П. Скулачева,
доказавшего с помощью этого объекта справедливость
представлений о трансмембранном протонном градиенте
как предшественнике АТФ.
Трансгенные растения с генами фенилпропаноидного и
терпеноидного метаболизма.
Как известно, в результате
включения сигнальных систем образуются растительные ан-
тибиотики - фитоалексины, относящиеся главным образом
к соединениям фенилпропаноидного и терпеноидного мета-
болизма клеток. Имеется несколько удачных попыток повы-
шения устойчивости растений с помощью привнесения генов
ферментов синтеза фенилпропаноидных [Hain et al., 1993;
Maher et al., 1994; Oommenet et al., 1994; Colliver et al., 1997;
Coutos-Thevenot et al., 2001; Jones et al.,
2001] и терпеноидных
[Kunkel et al., 1999; Ouwerkerk et al.,
1999] фитоалексинов.
К этому направлению работ примыкает усиление лигнифи-
кации клеток с помощью гена пероксидазы [Dowd et al., 1998;
Ostergaard et al.,
2000], катализирующей превращение моно-
мерных производных фенилпропаноидного метаболизма
(фенольных кислот и спиртов) в гетерополимер лигнин.