ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 25.10.2023
Просмотров: 88
Скачиваний: 2
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Адам мен жануарлардың денесінде простагландиндер мен тромбоксандар жалпы прекурсордан — көміртегі атомдарының тиісті саны бар маңызды полиқанықпаған май қышқылдарынан және молекулалардағы Қос байланыстардан, соның ішінде линол және арахидон қышқылдарынан түзіледі. Простагладин биосинтезінің жылдамдығын шектейтін фактор бос май қышқылдарының жалпы саны (пул) болып табылады, сондықтан құрамында полиқанықпаған май қышқылдары бар триглицеридтердің, фосфолипидтердің және холестерин эфирлерінің гидролитикалық ыдырауына әсер ететін заттар простагландиннің түзілу қарқындылығын реттей алады. Сонымен, катехоламиндер, брадикинин, ангиотензин II денеде май қышқылдарының бөлінуінің жоғарылауын тудырады, осылайша простагландиндердің түзілуін жанама түрде ынталандырады. Ишемия немесе жасушаларға механикалық әсер ету кезінде простагландиндердің, тромбоксандардың биосинтезін ынталандыру механизмі сияқты. Кортикостероидты гормондар, керісінше, простагландин мен тромбоксандардың биосинтезін тежейді, өйткені олар май қышқылдарының бөлінуін тежейді. Кейбір қосылыстар простагландиндер мен тромбоксандардың жекелеген түрлерінің түзілуіне әсер етеді, мысалы май қышқылының пероксиді простагландин I2-(простагландин I2 немесе простациклин) биосинтезін, ал имидазол — тромбоксан А2 түзілуін арнайы тежейді. Бірқатар дәрі-дәрмектер простагландиндер мен тромбоксандардың пайда болуына айқын әсер етеді, олардың жалпы санын ғана емес, сонымен қатар жекелеген түрлер мен сериялар арасындағы байланысты өзгертеді. мысалы, қабынуға қарсы әсері бар дәрілер — салицилаттар, индометацин (метиндол), бруфен және т. б. - простландин биосинтезінің бірінші сатысын катализдейтін циклооксигеназаны тежейді. Бұл простагландиндер мен тромбоксандардың түзілуінің төмендеуіне әкеледі.
Простагландиндер мен тромбоксандар қысқа мерзімді қосылыстар болып табылады. Олардың кейбіреулерінің жартылай өмір сүру уақыты секундтармен есептеледі. Простагландиндердің тез жойылуы олардың әсерлерінің локализациясын анықтайды-простагландин негізінен олардың синтезі орнында әрекет етеді. Простагландин метаболизмі олардың тез инактивациясына әкеледі, барлық тіндерде жүзеге асырылады, бірақ әсіресе өкпеде, бауырда және бүйректе белсенді.
Простагландиннің биологиялық әсері жеке простагландиндердің биологиялық поливаленттілігінің ғана емес, сонымен қатар олардың алуан түрлілігінің арқасында әр түрлі. F1 және D2 простагландиндері бронхтардың жиырылуын тудырады, ал простагландин Е2 олардың босаңсуын тудырады. Тромбоксан А2 қан тамырларының қабырғаларын жиырады және қан қысымын жоғарылатады, ал простагландин I2 гипотензивті әсермен бірге вазодилататор әсеріне ие. Тромбоксан А2 мен простагландин I2 арасындағы антагонистік қатынастар олардың қан ұю жүйесіне әсер етуімен де көрінеді: тромбоксан А2 тромбоциттер агрегациясының күшті табиғи индукторы болып табылады, ал қан тамырларының қабырғаларында синтезделген простагландин I2 адам мен жануарлар ағзасында тромбоциттер агрегациясының ингибиторы рөлін атқарады. Простагландин I2 мен тромбоксан А2 арақатынасы жүрек-қан тамырлары жүйесінің қалыпты жұмыс істеуі үшін өте маңызды.
Простагландиндер овуляция процесі үшін өте маңызды; олар жұмыртқаның алға жылжуына және сперматозоидтардың қозғалғыштығына, жатырдың жиырылу белсенділігіне әсер етеді, сонымен қатар қалыпты босану үшін қажет: әлсіз босану және жүктіліктің артуы P жетіспеушілігімен байланысты, ал P. түзілуінің жоғарылауы өздігінен түсік түсіруге және мерзімінен бұрын босануға әкелуі мүмкін. Жаңа туылған нәрестелерде P. кіндік тамырлары мен артериялық каналдың жабылуын реттейді.
Простагландиндер белгілі бір рецепторларға әсер етуден басқа, жасушаның функционалды құрылымдарына тікелей әсер ете алады. Дәрілік заттар ретінде простагландиндер босануды тудыру, босану белсенділігін қоздыру және ынталандыру, жүктілікті тоқтату үшін қолданылады. Терапевтік дозаларда простагландиндер ана мен ұрыққа теріс әсер етпейді. Жатырдың простагландинді енгізуге сезімталдығы жүктіліктің әртүрлі кезеңдерінде әр түрлі болады; өте ерте және кеш кезеңдерде ынталандырушы әсер оңай туындайды, ал олардың арасында простагландин препараттарын енгізуге миометрия әлсіз әсер етеді. Простагландинді аборт жасау, босану белсенділігін қоздыру және ынталандыру үшін қолдануға қарсы көрсеткіштер-ауыр соматикалық аурулар, простагландин препараттарына аллергиялық реакциялар, бронх демікпесі, эпилепсия, жатырдағы тыртық.
2.Бөлім
2.1 Гормон секрециясын реттеу
Гормоналды реттеу, қанға түсетін гормондардың қатысуымен жүзеге асырылатын жануарлар мен адам ағзасының өмірлік белсенділігін реттеу; функцияларды реттеу мен үйлестірудің жүйке және гуморальдық жүйелерімен тығыз байланысты функцияларды өзін-өзі реттеу жүйелерінің бірі. Маңызды биологиялық процестердің бірі-гормондардың бөлінуін реттеу, олардың түзілуін, жасушалардан бөлінуін және метаболизм процестерін және тіндер мен мүшелердің басқа функцияларын қолдау үшін қажетті мөлшерде айналымға түсуін қамтамасыз ету. Бұл реттеуші жүйенің құрамдас бөліктері гуморальдық факторлар болып табылады, оларға метаболизм өнімдері мен гормондар, нейро-гормоналды және жүйке факторлары жатады.
Метаболизм өнімдерінің гормон секрециясының әртүрлі кезеңдеріне әсерінің бірқатар мысалдарын келтіруге болады. Сонымен, гуморальды реттеулердің мысалы-ұйқы безі аралдарының бета-жасушаларынан инсулиннің жасушадан тыс кеңістікке және айналымға бөлінуі, гликемия деңгейінің жоғарылауымен аминқышқылдары да осы секрецияның тимуляторлары болып табылады, инсулиннің бөліну процесімен үйлеседі.оның биосинтезінің жоғарылауы байқалады. Қандағы қант деңгейінің төмендеуі инсулин секрециясының төмендеуіне, секрецияның жоғарылауына және оның гормоналды антагонистерінің — ұйқы безі аралдарының альфа жасушалары шығаратын глюкагонның, өсу гормонының, гидрокортизонның, адреналиннің және норадреналин медиаторының айналымына енуіне ықпал етеді. Күрделі метаболикалық процестердің нәтижесінде бірқатар гормондардың қатаң үйлестірілген өзара әрекеттесуі қандағы қант пен глюкоза метаболизмінің физиологиялық деңгейінің сақталуын қамтамасыз етеді. Оларға деген сұраныстың жоғарылауына жауап ретінде гормондардың секрециясын реттеуден басқа, олардың ақуыздармен байланысынан гормондардың бөлінуі маңызды. Қан плазмасында инсулин, тироксин, өсу гормоны, прогестерон, гидрокортизон, кортикостероп және басқа гормондарды байланыстыратын арнайы ақуыздар зерттелді. Гормондар мен ақуыздар салыстырмалы түрде төмен энергияға ие ковалентті емес байланыстармен байланысады, сондықтан бұл кешендер гормонды босату үшін оңай бұзылады. Ақуыздармен кешендеу гормонның бір бөлігін белсенді емес күйде ұстауға мүмкіндік береді. Сонымен қатар, бұл байланыс гормонды химиялық және энзи-матикалық факторлардың әсерінен қорғайды. Ақуыздармен байланысты гормондар айналымдағы көлік түрлерінің бірі болып табылады және олардың сақталуын қамтамасыз етеді деген түсінікке басқа фактілер қосылды: бұл кешендердің биологиялық маңыздылығының маңызды құрамдас бөлігі олардан бос, яғни белсенді, гормондарды тез босату мүмкіндігі болып табылады. Гормондардың секрециясын реттеу бірнеше байланысты механизмдер арқылы жүзеге асырылады. Оларды бүйрек үсті безінің негізгі глюкокортикоидты гормоны кортизол мысалында көрсетуге болады. Оның өнімдері гипоталамус деңгейінде әрекет ететін кері байланыс механизмі арқылы реттеледі. Қандағы кортизол деңгейі төмендеген кезде гипоталамус кортиколиберин бөледі, бұл гипофиздің кортикотропин (ACTH) секрециясын ынталандыратын фактор. ACTH деңгейінің жоғарылауы өз кезегінде бүйрек үсті безінде кортизол секрециясын ынталандырады, нәтижесінде қандағы кортизол мөлшері артады. Кортизол деңгейінің жоғарылауы кері байланыс механизмі бойынша кортиколибериннің бөлінуін тежейді-және қандағы кортизол мөлшері қайтадан төмендейді. Кортизол секрециясы тек кері байланыс механизмімен реттелмейді. Мәселен, мысалы, стресс кортиколибериннің босатылуын тудырады, сәйкесінше кортизол секрециясын арттыратын реакциялардың барлық сериясы. Сонымен қатар, кортизол секрециясы күнделіктімуаққа бағынады; ол оянғанда өте жоғары, бірақ ұйқы кезінде біртіндеп минималды деңгейге дейін төмендейді. Бақылау механизмдеріне гормонның метаболизм жылдамдығы және олардың белсенділігін жоғалту жатады. Ұқсас реттеу жүйелері басқа гормондарға да әсер етеді. Орталық жүйке жүйесі гормондардың секрециясын реттеуде маңызды. Эндокриндік бездердің қызметін үйлестіретін және бақылайтын ОЖЖ-нің маңызды бағыттарының бірі
гипоталамус болып табылады, онда аденогипофиз гормондарының синтезі мен секрециясын реттеуге қатысатын нейросекреторлық ядролар мен орталықтар локализацияланған. Гипоталамус-гипофизді реттеу кері байланыс принципі бойынша жұмыс істейтін механизмдер арқылы жүзеге асырылады, онда өзара әрекеттесудің әртүрлі деңгейлері айқын ерекшеленеді (сурет. 1).
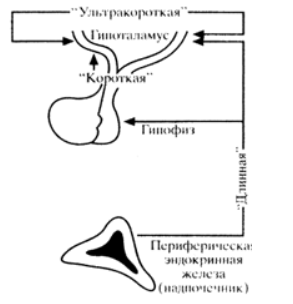
Сурет 2. Кері байланыс деңгейлері.
"Ұзын" кері байланыс тізбегі перифериялық эндокриндік бездің гипофиз және гипоталамус орталықтарымен (ОЖЖ-нің супрагипоталамикалық және басқа аймақтарымен) айналымдағы қандағы гормондардың өзгеретін концентрациясының аталған орталықтарына әсер ету арқылы өзара әрекеттесуін білдіреді."Қысқа" кері байланыс тізбегі гипофиздің тропиктік гормонының жоғарылауы (мысалы, ACTH) гипофизотропты гормонның (бұл жағдайда кортиколиберин) секрециясы мен бөлінуін модуляциялайтын және өзгертетін өзара әрекеттесуді білдіреді."Ультра қысқа" кері байланыс тізбегі-бір гипофизотропты гормонның бөлінуі басқа гипофизотропты гормонның бөлінуі мен бөліну процестеріне әсер ететін гипоталамус ішіндегі өзара әрекеттесу түрі. Кері байланыстың бұл түрі кез-келген эндокриндік безде болады. Сонымен, осы нейрондардың аксондары арқылы және жасушааралық өзара әрекеттесу арқылы (жасушадан жасушаға) окситоцин немесе вазопрессиннің бөлінуі осы гормондарды шығаратын нейрондардың белсенділігін өзгертеді. Тағы бір мысал, пролактиннің бөлінуі және оның васкулярлық кеңістіктерге диффузиясы көршілес лактотрофтарға әсер етеді, содан кейін пролактин секрециясының тежелуі."Ұзын "және" қысқа "кері байланыс тізбектері" жабық " типті жүйелер ретінде жұмыс істейді, яғни олар өзін-өзі реттейтін жүйелер. Алайда, олар ішкі және сыртқы сигналдарға жауап береді, қысқа уақытқа өзін-өзі реттеу принципін өзгертеді (мысалы, стресс кезінде және т.б.). Сонымен қатар, бұл жүйелерге күн мен түннің өзгеруіне байланысты биологиялық циркадиялық ритақты қолдайтын механизмдер әсер етеді. Циркадиялық ритақ организмнің гомеостазын реттейтін және қоршаған ортаның өзгеретін жағдайларына бейімделуге мүмкіндік беретін жүйенің құрамдас бөлігі болып табылады. Күндізгі-түнгімеақ туралы ақпарат ОЖЖ – ге көз торынан супрахиазматикалық ядроларға беріледі, олар эпифизбен бірге орталық циркадиялық механизмді - "биологиялық сағатты"құрайды. Күн-түн механизмінен басқа, осы "сағаттардың" қызметіне басқа реттеушілер қатысады (Дене температурасының өзгеруі, демалу, ұйқы және т.б.).
2.2. Көмірсу, липид, ақуыз және су - тұз алмасуының гормоналды реттелуі
Тірі организмнің негізгі энергетикалық ресурстары — көмірсулар мен майлар ферменттік катаболикалық түрлендірулер арқылы жасушаларда оңай алынатын потенциалдық энергияның жоғары қорына ие. Көмірсулар мен май алмасуларының, сондай-ақ гликолиз өнімдерінің биологиялық тотығу процесінде бөлінетін Энергия негізінен синтезделген АТФ фосфат байланыстарының химиялық энергиясына айналады. АТФ-да жинақталған макроэргиялық байланыстардың химиялық энергиясы, өз кезегінде, әртүрлі жасушалық жұмыстарға жұмсалады — электрохимиялық градиенттерді құру және қолдау, бұлшықеттердің жиырылуы, секреторлық және кейбір көлік процестері, ақуыз, май қышқылдарының биосинтезі және т. б. "отын" функциясынан басқа, көмірсулар мен майлар ақуыздармен бірге маңызды құрылыс, пластикалық жеткізушілер рөлін атқарады. жасушаның негізгі құрылымдарына кіретін материалдар - нуклеин қышқылдары, қарапайым ақуыздар, гликопротеидтер, бірқатар липидтер және т. б. Көмірсулар мен майлардың ыдырауына байланысты синтезделген АТФ жасушаларды жұмыс істеуге қажетті энергиямен қамтамасыз етіп қана қоймайды, сонымен қатар camp түзілу көзі болып табылады, сонымен қатар көптеген ферменттердің белсенділігін, құрылымдық ақуыздардың күйін реттеуге қатысады, олардың фосфорлануын қамтамасыз етеді. Жасушалар тікелей тастайтын көмірсулар мен липидті субстраттар-моносахаридтер (ең алдымен глюкоза) және эфирленбеген май қышқылдары (HEFA), сондай-ақ кейбір тіндерде кетон денелері. Олардың көздері-көмірсулардың гликоген түрінде және бейтарап майлар түрінде ағзаларға салынған ішектен сіңетін Тамақ өнімдері липидтер, сондай-ақ көмірсулар түзетін аминқышқылдары мен глицерин (глюконеогенез). Омыртқалылардағы депозиттік органдарға бауыр мен майлы (адипозды) тін, глюконеогенез органдарына — бауыр мен бүйрек жатады. Жәндіктерде май ағзасы депозиттік орган болып табылады. Сонымен қатар, глюкоза мен HEHA көздері жұмыс істейтін жасушада сақталған немесе өндірілген кейбір қосалқы немесе басқа өнімдер болуы мүмкін. Көмірсулар мен май алмасуының әртүрлі жолдары мен кезеңдері көптеген өзара әсерлермен байланысты. Бұл алмасу процестерінің бағыты мен қарқындылығы бірқатар сыртқы және ішкі факторларға байланысты. Оларға, атап айтқанда, тұтынылатын тағамның мөлшері мен сапасы және оның ағзаға ену ритағы, бұлшықет және жүйке қызметінің деңгейі және т. б. Жануар ағзасы үйлестіру механизмдерінің күрделі кешені арқылы тамақтану режимінің сипатына, жүйке немесе бұлшықет жүктемесіне бейімделеді. Сонымен, көмірсулар мен липидтер алмасуының әртүрлі реакцияларының барысын бақылау жасуша деңгейінде тиісті субстраттар мен ферменттердің концентрациясымен, сондай-ақ белгілі бір реакция өнімдерінің жинақталу дәрежесімен жүзеге асырылады. Бұл бақылау механизмдері өзін-өзі реттеу механизмдеріне жатады және бір жасушалы және көп жасушалы организмдерде жүзеге асырылады. Соңғысында көмірсулар мен майларды жоюдың реттелуі жасушааралық өзара әрекеттесу деңгейінде болуы мүмкін. Атап айтқанда, метаболизмнің екі түрі де өзара бақыланады: бұлшық еттердегі ЭЖК глюкозаның ыдырауын тежейді, ал май тініндегі глюкозаның ыдырау өнімдері ЭЖК түзілуін тежейді. Ең жоғары ұйымдастырылған жануарларда бүкіл организмнің метаболикалық процестерін бақылауда маңызды болып табылатын эндокриндік жүйенің эволюциясы процесінде пайда болуымен анықталатын аралық алмасуды реттеудің арнайы жасушааралық механизмі пайда болады. Омыртқалы жануарлардағы май мен көмірсулар алмасуын реттеуге қатысатын гормондардың ішінде келесілер орталық орын алады: тағамның қорытылуын және ас қорыту өнімдерінің қанға сіңуін бақылайтын асқазан-ішек гормондары; инсулин және глюкагон — көмірсулар мен липидтердің аралық алмасуының арнайы реттегіштері; СТГ және онымен функционалды байланысты "соматомединдер" және СИФ,глюкокортикоидтар, ACTH және адреналин спецификалық емес бейімделу факторлары болып табылады.