ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 23.11.2023
Просмотров: 243
Скачиваний: 2
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
-
Эпидермис — слой клеток, которые защищают от вредного воздействия среды и излишнего испарения воды. Часто поверх эпидермиса лист покрыт защитным слоем восковидного происхождения (кутикулой). -
Мезофилл, или паренхима — внутренняя хлорофиллоносная ткань, выполняющая основную функцию — фотосинтез. -
Сеть жилок, образованных проводящими пучками, состоящими из сосудов и ситовидных трубок, для перемещения воды, растворённых солей, сахаров и механических элементов. -
Устьица — специальные комплексы клеток, расположенные, в основном, на нижней поверхности листьев; через них происходит испарение воды и газообмен.
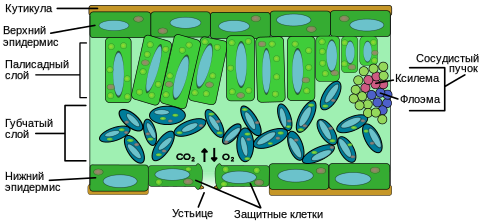
Эпидерма — наружный слой многослойной структуры клеток, покрывающий лист со всех сторон; пограничная область между листом и окружающей средой. Эпидерма выполняет несколько важных функций: защищает лист от излишнего испарения, регулирует газообмен с окружающей средой, выделяет вещества обмена и в некоторых случаях впитывает воду. Большинство листьев имеют дорсовентральную анатомию: верхняя и нижняя поверхности листа имеют различную структуру и выполняют разные функции.
Эпидерма обычно прозрачна (в её строении отсутствуют либо присутствуют в недостаточном количестве хлоропласты) и снаружи покрыта защитным слоем восковидного происхождения (кутикула), который препятствует испарению. Кутикула нижней части листа, как правило, тоньше, чем на верхней, и толще в биотопах с засушливым климатом по сравнению с теми биотопами, где недостаток влаги не ощущается.
В состав ткани эпидермы входят следующие типы клеток: эпидермальные (или двигательные) клетки, защитные клетки, вспомогательные клетки и трихомы. Эпидермальные клетки самые многочисленные, крупные и наименее приспособленные. У однодольных растений они более растянуты, чем у двудольных. Эпидерма покрыта порами, называемыми устьицами, которые являются частью целого комплекса, состоящего из поры, со всех сторон окружённой содержащими хлоропласт защитными клетками, и от двух до четырёх побочных клеток, в которых хлоропласт отсутствует. Этот комплекс регулирует испарение и газообмен листа с окружающей средой. Как правило, количество устьиц на нижней части листа больше, чем на верхней. У многих видов поверх эпидермиса вырастают трихомы.
Большую часть внутренности листа между верхним и нижним слоями эпидермиса составляет паренхима (основная ткань), или мезофилл. В норме мезофилл образован хлорофиллсинтезирующими клетками, поэтому употребляется и синонимичное название — хлоренхима. Продукт фотосинтеза называется фотосинтат.
У папоротников и большинства цветковых растений мезофилл разделён на два слоя:
-
Верхний, палисадный слой плотно упакованных, вертикально-расположенных клеток прямо под верхним слоем эпидермиса; толщиной в одну или две клетки. Клетки этого слоя содержат гораздо больше хлоропластов, чем в нижележащем губчатом слое. Длинные клетки цилиндрической формы, как правило, уложены в один — пять слоёв. Они, находясь близко к границе листа, расположены оптимально для получения солнечного света. Небольшие промежутки между клетками используются для поглощения углекислого газа. Промежутки должны быть достаточно малыми, чтобы поддерживать капиллярное действие по передаче воды. Растения должны адаптировать свою структуру для оптимального получения света при различных природных состояниях, таких как солнце или тень — солнечные листья имеют многослойный палисадный слой, в то время как теневые и старые, лежащие близко к земле листья имеют только один слой. -
Клетки нижнего, губчатого слоя упакованы рыхло и, вследствие этого, губчатая ткань обладает большой внутренней поверхностью благодаря развитой системе межклетников, сообщающихся друг с другом и с устьицами. Рыхлость губчатой ткани играет важную роль в газообмене листа кислородом, углекислым газом и парами воды.
Листья обычно окрашены в зелёный цвет благодаря хлорофиллу — фотосинтезирующему пигменту, находящемуся в хлоропластах — зелёных пластидах. Растения, у которых ощущается недостаток либо отсутствие хлорофилла, не могут фотосинтезировать.
В некоторых случаях в результате соматических мутаций возможно образование участков мезофилла мутантными клетками, не синтезирующими хлорофилл, при этом листья таких растений имеют пёструю окраску, обусловленную чередованием участков нормального и мутантного мезофилла.
Растения в умеренных и северных широтах, а также в сезонно-сухих климатических зонах могут быть листопадными, то есть их листья с приходом неблагоприятного сезона опадают либо отмирают. Этот механизм имеет название сбрасывания или опадания. На месте опавшего листа на веточке образуется рубец —
листовой след. В осенний период листья могут окраситься в жёлтый, оранжевый или красный цвет, так как с уменьшением солнечного света растение уменьшает выработку зелёного хлорофилла, и лист приобретает окраску вспомогательных пигментов, таких как каротиноиды и антоцианы.
Жилки листа являются сосудистой тканью и расположены в губчатом слое мезофилла. По рисунку разветвления жилки, как правило, повторяют структуру разветвления растения. Жилки состоят из ксилемы — ткани, служащей для проведения воды и растворённых в ней минеральных веществ, и флоэмы — ткани, служащей для проведения органических веществ, синтезируемых листьями. Обычно ксилема лежит поверх флоэмы. Вместе они образуют основную ткань, называемую сердцевиной листа.
Тема 4. Генеративные органы семенных растений
-
вопрос. Опишите строение семязачатка цветковых и его составных частей, приведите рисунок. Перечислите морфологические типы семязачатков у цветковых. Как происходит развитие семязачатка, в чем заключается имеющий место процесс мегаспорогенеза?
Строение семязачатков (семяпочек) цветковых растений во многом сходно со строением семязачатков голосеменных. Но в то время как у голосеменных преобладают в общем массивные семязачатки, обычно с толстыми, иногда даже грубыми покровами и со сравнительно большим запасом питательных веществ, у цветковых семязачатки обычно небольшие, часто очень мелкие, в большинстве случаев со слабо развитыми покровами и почти совершенно лишены запасов питательных веществ. Оказавшись в полости завязи, этой своеобразной влажной камере, семязачатки могут формироваться быстро и с минимальной затратой строительного материала, что оказалось исключительно важным биологическим преимуществом. Это обстоятельство особенно подчеркивал известный русский ботаник М. И. Голепкии (1927), который в книге «Победители в борьбе за существование» писал следующее: «Уменьшение затраты питательных и строительных веществ на семяпочки до оплодотворения только и позволило, весьма возможно, покрытосеменным развить необычайную, так сказать, деятельность в деле выработки приспособлений, с одной стороны, для защиты зародышей (в семенах и плодах), с другой — для расселения по поверхности земли. В отношении увеличения количества семян, органов размножения, стоит сравнить покрытосеменные опять-таки с голосеменными, например, какой-нибудь тополь с елью или сосной — у первого буквально неисчислимые количества семян, у вторых очень ограниченные. Про травянистые формы и говорить нечего». Очень хорошо сказано. Действительно, экономия материала, используемого на построение семязачатка, несомненно, позволила цветковым растениям существенно повысить семенную продуктивность, или урожайность. В то же время та же экономия материала привела не только к значительному упрощению семязачатка, но и к более быстрому его формированию, чем у голосеменных.
П
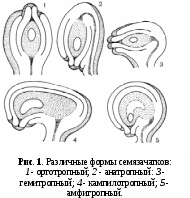 режде чем перейти к строению семязачатка, рассмотрим его основные морфологические типы (рис. 1), установленные еще французским ботаником Ш. Мирбезем в 1829 г.
режде чем перейти к строению семязачатка, рассмотрим его основные морфологические типы (рис. 1), установленные еще французским ботаником Ш. Мирбезем в 1829 г.У многих цветковых растений микропиле семязачатка находится на одной оси с семяножкой (фуникулусом) и, таким образом, семязачаток расположен перпендикулярно к плаценте. Такой радиально-симметричный семязачаток был назван ортотропным (от греч. orthos — прямой, правильный и tropos — поворот, направление) или прямым. (рис. 1). Нередко его называют атропным (от греч. а — частица отрицания). Ортотропные семязачатки характерны для хлорантовых, савруровых, перцевых, роголистника, платана, мириковых, ореховых, валлиснерии, взморника, рдеста, ксирисовых, эриокауловых, рестиевых и некоторых других родов и семейств.
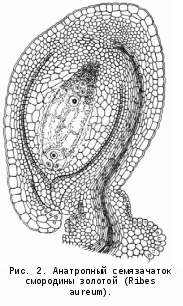
Гораздо более распространены семязачатки, загнутые вниз (в сторону плаценты) на 180° и приросшие вследствие этого своим боком к удлиненной семяножке. Поэтому микропиле у них обращено к плаценте и расположено рядом с основанием семязачатка. Такие семязачатки называются анатропными (от греч. anatrope — перевернутый, обращенный) или обращенными (рис. 1, 2). Анатропные семязачатки характерны для большинства цветковых, как двудольных, так и однодольных. Известный чешский ботаник Ф. Нетолицкий (1926) пришел к выводу, что анатропный тип является исходным, а ортотропный — производным, что вполне согласуется с современными представлениями.
Наряду с анатропными и ортотропными семязачатками существует еще несколько их «архитектурных» типов, которые, однако, меньше распространены.. Следует прежде всего упомянуть гемитропные (от греч. hemi — половина, или, точнее, «полу»), или полуобращенные, семязачатки, повернутые только на 90° по отношению к плаценте. Они характерны, например, для казуарины, некоторых первоцветных, некоторых бурачниковых, губоцветных, пасленовых и норичниковых, мальпигиевых, некоторых лилейных и ряда других растений. Гемитропные семязачатки представляют собой промежуточный тип между анатропными и ортотропными (рис. 1).
В
 тех случаях, когда семязачаток вследствие более сильного разрастания одной стороны становится изогнутым, он называется кампилотропным (от греч. kampylos — изогнутый) или полусогнутым. В отличие от анатропного и гемитропного типов здесь изогнуты интегумент и даже мегаспорангий (нуцеллус). Этот тип семязачатка характерен для многих представителей порядка гвоздичных, для каперсовых, некоторых бобовых, для филезиевых, некоторых злаков и ряда других групп. В большинстве случаев кампилотропные семязачатки возникают из анатропных, но не исключено, что иногда они могут возникать и из ортотропных.
тех случаях, когда семязачаток вследствие более сильного разрастания одной стороны становится изогнутым, он называется кампилотропным (от греч. kampylos — изогнутый) или полусогнутым. В отличие от анатропного и гемитропного типов здесь изогнуты интегумент и даже мегаспорангий (нуцеллус). Этот тип семязачатка характерен для многих представителей порядка гвоздичных, для каперсовых, некоторых бобовых, для филезиевых, некоторых злаков и ряда других групп. В большинстве случаев кампилотропные семязачатки возникают из анатропных, но не исключено, что иногда они могут возникать и из ортотропных.
В некоторых случаях семязачаток перегибается посередине таким образом, что в продольном разрезе его мегаспорангий имеет подковообразную форму. Это амфитропный (от греч. amphi — оба) семязачаток. Он встречается у лейтнерии, кроссосомы, кнеоровых некоторых акантовых и немногих других растений (рис. 2).
Между всеми этими типами имеются промежуточные формы, и поэтому очень часто бывает трудно определить, какой тип семязачатка имеет то или иное растение. Тем не менее выделение этих типов оказалось очень полезным, так как они имеют определенное систематическое значение.
К
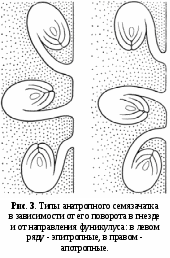 ак это уже давно указывал датский ботаник Й. Э. Варминг (1913), в систематическом отношении важно также, куда загнуты анатропные или кампилотропные семязачатки — к верхушке (в этом случае они называются эпитропными) или ко дну завязи (они носят название апотропных; рис. 3).
ак это уже давно указывал датский ботаник Й. Э. Варминг (1913), в систематическом отношении важно также, куда загнуты анатропные или кампилотропные семязачатки — к верхушке (в этом случае они называются эпитропными) или ко дну завязи (они носят название апотропных; рис. 3).Основные черты строения семязачатков мы уже рассматривали в предыдущем томе «Жизни растений». Но напомним еще раз, что семязачаток состоит из двух основных частей — мегаспорангия (нуцеллуса) и окружающего его покрова, называемого интегументом. Интегумент не полностью закрывает мегаспорангий, на верхушке семязачатка его края не срастаются полностью и оставляют отверстие в виде канала для внедрения пыльцевых трубок. Это микропиле, или, иначе, семявход. Нижняя часть семязачатка, т. е. место ее перехода в семяножку (фуникулус), называется халазой. Некоторые авторы, в том числе даже такой известный эмбриолог, как индийский ботаник П. Махешвари (1950), весь семязачаток приравнивают к мегаспорангию. Это большая ошибка. Семязачаток — это не только мегаспорангий, это мегаспорангий плюс интегумент.
Читатель может спросить: почему мегаспорангий семязачатка называют нуцеллусом, если известно, что это мегаспорангий? Дело в том, что долгое время это не было известно, пока в 1851 г. великий немецкий ботаник В. Гофмейстер не установил, что образование, давно известное под названием нуцеллуса, представляет собой не что иное, как мегаспорангий. Но, как часто бывает в науке, даже после замечательных исследований Гофмейстера мегаспорангий семенных растений продолжали называть нуцеллусом, а некоторые ботаники, как, например, американский морфолог А. Имс (1961), подвергали сомнению даже само наличие мегаспорангия у цветковых растений.