Файл: Вопросы к экзамену патология как наука. Место патологии в фармации и среди других медикобиологических наук.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 09.01.2024
Просмотров: 457
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
Увеличение содержания натрия в организме может быть результатом общего нарушения обмена веществ. Такие состояния, как лихорадка, белковое голодание, сопровождаются задержкой натрия и воды в организме.
Последствия гипернатриемии сводятся прежде всего к повышению осмотического давления крови и внеклеточной жидкости, в результате чего внутриклеточная жидкость переходит во внеклеточное пространство. Возникает дегидратация клеток и их сморщивание, нарушение функции.
Избыток натрия во внеклеточной жидкости способствует задержке воды и развитию отеков (см. § 217), а также развитию гипертонии, так как натрий потенцирует действие адреналина на гладкую мускулатуру артериол и способствует их сужению. Известно, например, что у лиц с высоким суточным потреблением поваренной соли (30—35 г/сут) артериальное давление выше, чем у людей, потребляющих не более 5— 8 г соли.
Кальций и магний находятся преимущественно в костях в форме фосфорнокислых и отчасти углекислых и фтористых солей. Кальций в костной ткани образует нерастворимые соединения типа оксиапатита Са10(НР04)6(ОН)2, составляющие основу кристаллической структуры обызвествленных тканей (костей и твердых тканей зубов: эмали, дентина и цемента).
Помимо костной ткани, кальций и магний в небольшом количестве входят также в состав всех других клеток и тканей и биологических жидкостей. Содержание кальция в плазме крови составляет 2,5 ммол/л
Гипокальциемия — уменьшение содержания кальция в крови ниже 2 ммоль/л может быть результатом недостаточного поступления кальция с пищей, нарушения всасывания ионизированного кальция в кишечнике и нарушения функции ряда желез внутренней секреции — паращитовидных, щитовидной, надпочечников и поджелудочной железы.
Потребность взрослых людей в кальции составляет около 8 мг/кг массы. У беременных и кормящих матерей эта потребность возрастает до 24 мг/кг, у грудных детей потребность в кальции равна 50—55 мг/кг. Количество кальция, которое взрослый или детский организм должен получить с пищей для того, чтобы удовлетворить указанные потребности, в очень сильной степени зависит от характера пищи и от содержания в ней различных соединений, способствующих или тормозящих всасывание кальция в кишечнике
Всасывание кальция затрудняется при значительном избытке в пище фосфора, в силу чего образуется труднорастворимый трехосновной фосфорнокислый кальций (оптимальное соотношение Са/Р составляет 1 : 1,3—1,5). Такое же влияние на всасывание кальция оказывает и избыток в пище жиров, когда образуются почти нерастворимые соли кальция с жирными кислотами (кальциевые мыла). Заметно ухудшается всасывание кальция при наличии в пище значительных количеств щавелевой кислоты и фитина (гексафосфатинозита), также в результате образования нерастворимых солей кальция.
Следующим важным фактором, ограничивающим всасывание кальция в тонком кишечнике является недостаток витамина D.
Гормон коры надпочечников — кортизол способствует значительному повышению выделения кальция как с мочой, так и через кишечник, так как при этом тормозится всасывание кальция в кишечной стенке и реабсорбция его в канальцах почек.
Вследствие стойкой гипокальциемии изменяется нервно-мышечная возбудимость и сократимость мышц. В физиологических условиях ионы кальция снижают проницаемость клеточной мембраны для ионов. При недостатке кальция во внеклеточной жидкости проницаемость клеточной мембраны возрастает и ионы перемещаются по градиенту концентрации, мембранный потенциал падает, в мышечной клетке возникает спонтанное сокращение. Этому способствует и поступление в клетку свободного кальция. Последний активирует АТФ-азу мышечной клетки и способствует расщеплению АТФ с освобождением необходимой для сокращения мышцы энергии. По такому механизму возникают приступы спонтанных мышечных сокращений — тетания при гипофункции паращитовидных желез или при удалении их у животных в эксперименте.
Гиперкальциемия — повышение уровня кальция в сыворотке крови выше 2,5—3 ммоль/л. Наиболее важным фактором, ведущим к гиперкальциемии является гиперфункция паращитовидных желез — гиперпаратиреоз. Избыток паратгормона увеличивает дифференциацию стволовых клеток в остеокласты, а также увеличивает активность каждого остеокласта; тормозит дифференциацию остеокластов в остеобласты и тем самым снижает количество последних и, наконец, снижает активность каждого остеобласта. В результате костная ткань теряет кальций. Костная ткань заменяется фиброзной, становится мягкой — возникает фиброзная остеодистрофия. Количество кальция в крови при этом повышается, концентрация неорганического фосфора снижается. Этому способствует и усиленное всасывание кальция в кишечнике и реабсорбция в почках. В почках происходит обызвествление клеток канальцевого эпителия и выпадение фосфорнокислых и углекислых солей кальция в просвете канальцев. Иногда это является основой для образования камней в мочевом тракте.
В какой-то мере подобные явления могут возникать и при избытке в организме витамина D, который в больших дозах имитирует эффекты паратгормона.
Относительная гиперкальциемия может возникать при ацидозе, когда кальций переходит из неактивной белковосвязанной формы в ионизированную— активную форму.
Длительная гиперкальциемия может привести к снижению нервно-мышечной возбудимости, появлению парезов, параличей. На ЭКГ удлиняется интервал S — Т.
Основные принипы терапии:
устранение причин, вызвавших гипер или гипо содержание иона
назначение солевых растворов при гипо
проведение мероприятий ослебляющих гемодилюцию ( ограничение поступления воды в организм) ри гипо
введение средств активизирующих процессы выведения иона через почки ( диуретики) и через кишечник ( катионообменные смолы)
-
Виды и механизмы развития ацидоза и алкалоза. Газовый и негазовый ацидоз, алкалоз. Роль бикарбонатного буфера.
Буферные системы крови
Установлено, что состоянию нормы соответствует определенный диапазон колебаний рН крови — от 7,37 до 7,44 со средней величиной 7,40. Кровь представляет собой взвесь клеток в жидкой среде, поэтому ее кислотно-основное равновесие поддерживается совместным участием буферных систем плазмы и клеток крови. Важнейшими буферными системами крови являются бикарбонатная, фосфатная, белковая и наиболее мощная гемогло-биновая.
Бикарбонатная буферная система—мощная и, пожалуй, самая управляемая система внеклеточной жидкости и крови. На долю бикарбонатного буфера приходится около 10% всей буферной емкости крови. Бикарбонат
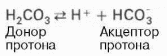
Для данной буферной системы величину рН в растворе можно выразить через константу диссоциации угольной кислоты (рКН2СО3) и логарифм концентрации недиссоциированных молекул Н2СО3 и ионов HCO3—:
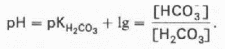
Ная система представляет собой сопряженную кислотно-основную пару, состоящую из молекулы угольной кислоты Н2СО3, выполняющую роль донора протона, и бикарбонат-иона НСО3—, выполняющего роль акцептора протона:
Истинная концентрация недиссоциированных молекул Н2СО3 в крови незначительна и находится в прямой зависимости от концентрации растворенного углекислого газа (СО2 + Н2О <=> Н2СО3). Поэтому удобнее пользоваться тем вариантом уравнения, в котором рКН2СО3 заменена «кажущейся» константой диссоциации Н2СО3, учитывающей общую концентрацию растворенного СО2 в крови:
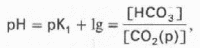
Где K1 — «кажущаяся» константа диссоциации Н2СО3; [СО2(р)] — концентрация растворенного СО2.
При нормальном значении рН крови (7,4) концентрация ионов бикарбоната НСО3 в плазме крови превышает концентрацию СО2 примерно в 20 раз. Бикарбонатная буферная система функционирует как эффективный регулятор в области рН 7,4.
Механизм действия данной системы заключается в том, что при выделении в кровь относительно больших количеств кислых продуктов водородные ионы Н+ взаимодействуют с ионами бикарбоната НСО3—, что приводит к образованию слабодиссоциирующей угольной кислоты Н2СО3. Последующее снижение концентрации Н2СО3 достигается в результате ускоренного выделения СО2 через легкие в результате их гипервентиляции (напомним, что концентрация Н2СО3 в плазме крови определяется давлением СО2 в альвеолярной газовой смеси).
Если в крови увеличивается количество оснований, то они, взаимодействуя со слабой угольной кислотой, образуют ионы бикарбоната и воду. При этом не происходит сколько-нибудь заметных сдвигов в величине рН. Кроме того, для сохранения нормального соотношения между компонентами буферной системы в этом случае подключаются физиологические механизмы регуляции кислотно-основного равновесия: происходит задержка в плазме крови некоторого количества СО2 в результате гиповентиляции легких. Как будет показано далее, данная буферная система тесно связана с гемоглобиновой системой.
Фосфатная буферная система представляет собой сопряженную кислотно-основную пару, состоящую из иона Н2РО4— (донор протонов) и иона НРО42— (акцептор протонов):
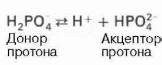
Роль кислоты в этой системе выполняет однозамещенный фосфат NaH2PO4, а роль соли двузамещенный фосфат—Na2HPO4.
Фосфатная буферная система составляет всего лишь 1% от буферной емкости крови. В других тканях эта система является одной из основных. Для фосфатной буферной системы справедливо следующее уравнение:
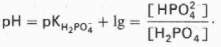
Во внеклеточной жидкости, в том числе в крови, соотношение [НРО42—]: [Н2РО4—] составляет 4:1. Величина рКН2РО4— равна 6,86.
Буферное действие фосфатной системы основано на возможности связывания водородных ионов ионами НРО42— с образованием Н2РО4— (Н+ + + НРО42— —> Н2РО4—), а также ионов ОН— с ионами Н2РО4— (ОН— + + Н2РО4——> HPO42— + H2O). Буферная пара (Н2РО4——НРО42—) способна оказывать влияние при изменениях рН в интервале от 6,1 до 7,7 и может обеспечивать определенную буферную емкость внутриклеточной жидкости, величина рН которой в пределах 6,9—7,4. В крови максимальная емкость фосфатного буфера проявляется вблизи значения рН 7,2. Фосфатный буфер в крови находится в тесном взаимодействии с бикарбонатной буферной системой. Органические фосфаты также обладают буферными свойствами, но мощность их слабее, чем неорганического фосфатного буфера.
Белковая буферная система имеет меньшее значение для поддержания КОР в плазме крови, чем другие буферные системы.
Белки образуют буферную систему благодаря наличию кислотно-основных групп в молекуле белков: белок—Н+ (кислота, донор протонов) и белок (сопряженное основание, акцептор протонов). Белковая буферная система плазмы крови эффективна в области значений рН 7,2—7,4.
Гемоглобиновая буферная система—самая мощная буферная система крови. Она в 9 раз мощнее бикарбонатного буфера; на ее долю приходится 75% от всей буферной емкости крови.
Участие гемоглобина в регуляции рН крови связано с его ролью в транспорте кислорода и углекислого газа. Константа диссоциации кислотных групп гемоглобина меняется в зависимости от его насыщения кислородом. При насыщении кислородом гемоглобин становится более сильной кислотой (ННЬО2). Гемоглобин, отдавая кислород, превращается в очень слабую органическую кислоту (ННЬ).
Итак, гемоглобиновая буферная система состоит из неионизированного гемоглобина ННЬ (слабая органическая кислота, донор протонов) и калиевой соли гемоглобина КНЬ (сопряженное основание, акцептор протонов). Точно так же может быть рассмотрена оксигемоглобиновая буферная система. Система гемоглобина и система оксигемоглобина являются вза-имопревращающимися системами и существуют как единое целое. Буферные свойства гемоглобина прежде всего обусловлены возможностью взаимодействия кисло реагирующих соединений с калиевой солью гемоглобина с образованием эквивалентного количества соответствующей калийной соли кислоты и свободного гемоглобина: