ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.05.2019
Просмотров: 15159
Скачиваний: 504

щества выносятся волнами в зону заплеска. Перекаты играют дру-
гую роль — здесь вспененный и «забуруненный» поток обогащает-
ся кислородом.
Третий механизм связан с вогнутыми берегами излучин и фор-
мирующимися здесь «приглубыми» местами — плесами и бочага-
ми (на самых малых водотоках). В плесах осуществляется связь реки
с горизонтом подземных вод; здесь «разгружаются» холодные род-
ники, поставляющие реке чистую воду. Кроме того, на дне плесов
происходит медленное разрушение различных органических ос-
татков до полной их минерализации за счет процессов брожения
с образованием метана, сероводорода и других продуктов бескис-
лородного процесса распада.
Наконец, четвертый и, пожалуй, самый важный механизм —
это зарастание реки водными растениями: плавающими, погру-
женными в воду и подводными, которые, в свою очередь, дают
прибежище многим сотням и тысячам живых организмов. Мел-
кие беспозвоночные, находящиеся в потоке, обычно живут на
поверхности растений или камней, следовательно, чем больше
водных растений, тем больше беспозвоночных, которые смогут
найти здесь пристанище. Часть из них действительно питается ра-
стениями, часть — разнообразными частицами, которые можно
подобрать тут же, в зоне «торможения» потока над подводными
зарослями. Растения могут выступать в роли покровителей, осо-
бенно для отдельного цикла развития животных (личинок), кото-
рые в противном случае были бы съедены или вымыты потоком.
Другие виды, такие как стрекозы, используют надводные части
растений, чтобы выбраться из воды, когда переходят из фазы
личинок к взрослому образу жизни. Увеличивая численность бес-
позвоночных, растения тем самым увеличивают объем потенци-
ального корма для рыб. Они образуют заросшие мелководья, в
которых молодь рыб и просто мелкая рыбешка спасаются от силь-
ного течения, хищников; вообще подводные заросли предостав-
ляют прекрасное убежище для размножения (метания икры, вы-
ведения мальков). Наконец, некоторые виды рыб просто питают-
ся растениями.
Все вместе: растения, животные и микроорганизмы — состав-
ляют единый речной биогеоценоз — аквальный комплекс. Водные
растения, или, как их еще называют, гидрофиты, — главные «на-
селенцы» наших рек, образующие основу биоты. Зарастание рек
проявляется в захвате (освоении) растительными сообществами
гидрофитов практически всех элементов русла, находящихся на
глубине не более 4 — 5 м, в увеличении числа и видового разнооб-
разия растительных группировок и в возрастании производимой
ими биомассы.
Функционирование речных аквальных комплексов определя-
ется взаимодействием речного потока, биоты и аллювиальных
136
русловых форм, причем в роли ведущего фактора дифференциа-
ции, развития и динамики русловых биоценозов выступает имен-
но речной поток.
Растениям непросто существовать в речных водах: как и любой
другой неподвижный объект, находящийся в движущейся среде,
водные растения оказывают сопротивление потоку и подвергаются
вследствие этого постоянному физическому воздействию: рас-
пластыванию, волочению, полосканию, истиранию; кроме того,
они могут быть просто сломаны потоком или вымыты из субстрата.
Разные виды растений по-разному переносят эти «сверхнагрузки»
водной среды, поэтому местообитания русла, различающиеся по
условиям течения, различаются и набором видов, слагающих
растительные сообщества, а также общим характером биоты реки.
Вдобавок скоростное поле во многом определяет состав и крупность
донных отложений (от тонких илов до крупной гальки), а
растения, в свою очередь, также проявляют избирательность к
тому или иному субстрату.
Но и биота, в свою очередь, трансформирует речную среду:
однажды появившись на поверхности речного ложа, растение из-
меняет модель течения, уменьшая скорость потока и увеличивает
его турбулентность, что впоследствии приводит к изменению ха-
рактера отложений и созданию условий, пригодных для обитания
других менее «экстремальных» видов.
Распространение биоты по дну руслового ложа может начаться
из одной точки или одновременно из нескольких, при этом за-
метное влияние группировок растительности на общую модель
течения становится ощутимым начиная с момента, когда биота
занимает 30 % площади ложа и около 10—15 % поперечного сече-
ния потока.
Наиболее узкими водными артериями являются реки 1-го
и 2-го порядков, русла которых могут быть затенены деревьями и
высокими травами, вследствие чего ложе бывает совершенно ли-
шено растительности. Заметно сказывается роль затенения и на
реках 3-го, 4-го порядков. В этом случае затененные травами пой-
мы прибрежные части русла бывают свободны от зарастания и
группировки водных растений развиваются на середине — в стреж-
невой части, что вызывает со временем перераспределение ско-
ростей и образование осередка. Большое влияние на характер за-
растания реки оказывает форма поперечного сечения руслового
ложа (рис. 4.11).
Наиболее широкие водные потоки — это, как правило, ниж-
ние участки рек. В широком потоке биота развивается на склонах и
отмелях речного ложа, где глубины невелики, а субстрат стаби-
лен. При этом из состава растительных сообществ выпадает целый
ряд видов, способных твердо укореняться только по берегам ру-
сел рек, но не в речном ложе.
137
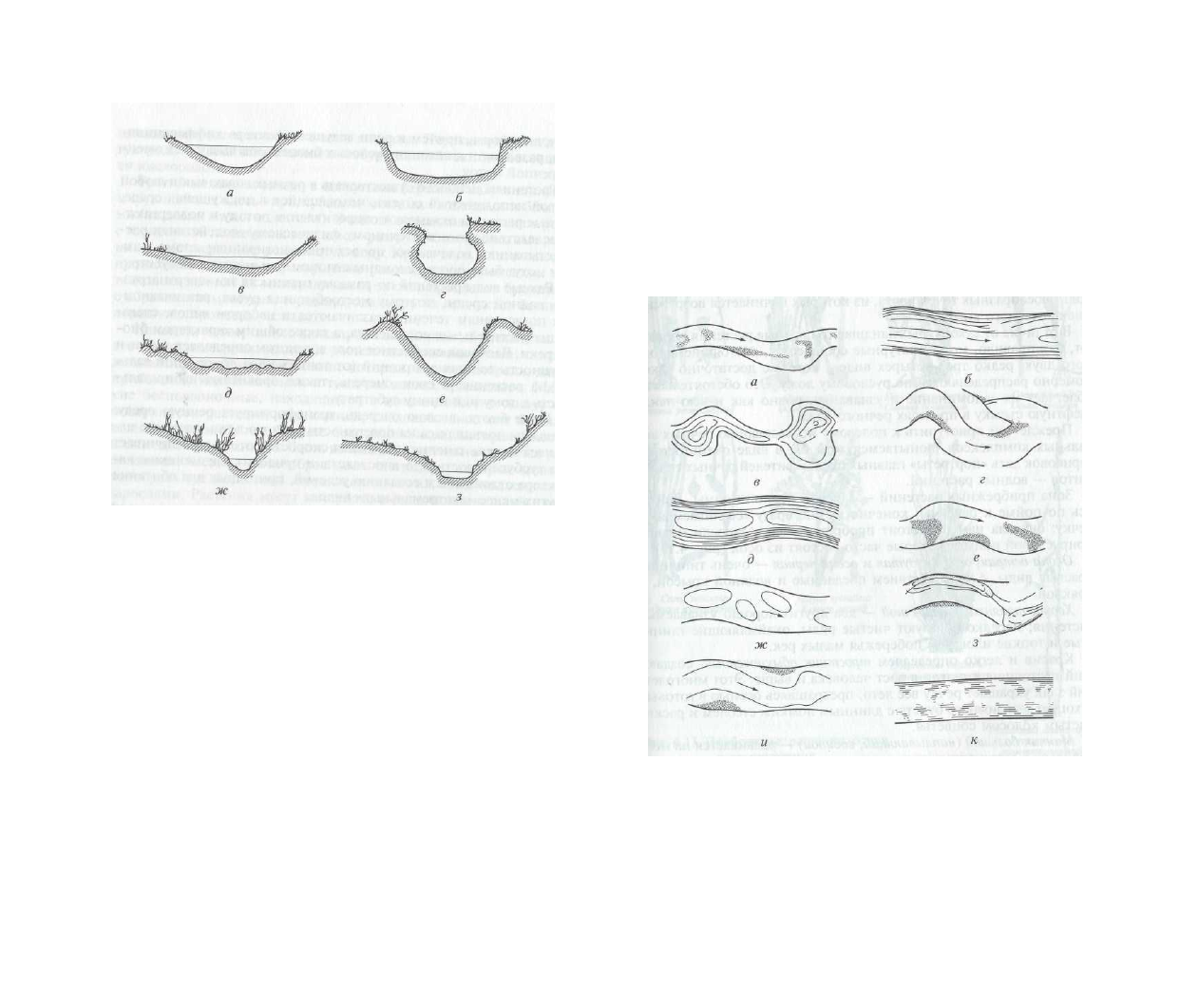
Рис. 4.11. Типы поперечного сечения заросших русел малых рек, опреде-
ляющие морфологию речных аквальных комплексов:
а — нормальное; б
— корытообразное; в — линзовидное; г — подковообразное; д — сложное; е —
V-образное; ж — канавообразное (канал-водоприемник); з — «поток»
(дренажная канава)
Глубиной потока определяется вероятность пересыхания и ко-
личество энергии, достигающей вегетативные органы растений.
Мелкие водотоки 1-го и 2-го порядков часто пересыхают в ме-
жень. Надводные гидрофиты могут перенести отсутствие воды в
русле, но плавающие и погруженные растения, подвергаясь пере-
сыханию, обычно отмирают.
Погруженные растения заметно угнетаются даже при простом
падении уровня воды. В среднем большая часть заселенных место-
обитаний малых рек имеет глубины до 1,5 м; для полного разви-
тия лишь некоторых видов (кубышка желтая, кувшинка чисто-
белая, лютик водный) необходима большая глубина — 3 —5 м. '
Таким образом, русловые процессы определяют характер и
структуру речных аквальных комплексов: конкретный набор рус-
ловых типов с набором местообитаний, заселение которых про-
исходит в соответствии с экологическими характеристиками ра-
стений в процессе взаимодействия зарождающейся биоты с вод-
138
ным потоком и наносами. Набор местообитаний и разнообразие
экологических условий последовательно изменяются от прямых и
слабоизогнутых русел к свободно меандрирующим и разветвлен-
ным, причем наиболее сложную структуру имеют аквальные ком-
плексы средних течений рек (рис. 4.12).
Аквальные комплексы — прекрасный объект для наблюдения,
поскольку они очень динамичны. Сезонные проявления неравно-
мерности стока (чередование аккумуляции и размыва, прохожде-
ние экстремальных половодных расходов) накладываются на изме-
нение состояний биоты, обусловленное процессами вегетации (раз-
Рис. 4.12. Разнообразие заросших русел малых рек — плановые очерта-
ния:
а — слабоизогнутое русло с небольшими песчаными грядами; 6 — корытообраз-
ное русло; в — четковидное русло с чередованием бочагов и перекатов; г —
свободно меандрирующее русло; д — углубленное русло; е — слабоизогнутое
русло с языковыми грядами; ж — прямое русло с хаотично расположенными
ямами; з — русло с ограниченной возможностью размыва берегов; и — подпор-
ный участок русла; к — искусственное русло (канал)
139

витие корневой системы и изменение силы сцепления с грунтом,
увеличение гидравлического сопротивления по мере разрастания
вегетативных частей растений). Одни виды быстрее поднимаются в
рост и уже к июню достигают максимального развития, другие ра-
стут медленнее, но, вырастая, затеняют группировки первых и со-
держание биоты претерпевает изменения. В результате с весны до
осени река почти каждую неделю заметно меняет свой облик.
К зиме большинство сообществ биоты реки отмирают. Остают-
ся лишь некоторые вечнозеленые виды (кубышка, омежник), со-
общества которых весной после прохождения половодья играют
роль своеобразных точек роста, из которых начинается возрожде-
ние аквального комплекса.
В отличие от сухопутных ландшафтов водные растения образу-
ют, как правило, монокультурные сообщества, состоящие из од-
ного-двух, редко трех-четырех видов, которые достаточно зако-
номерно распределяются по русловому ложу. Это обстоятельство
облегчает их запоминание и узнавание, ровно как и всю ланд-
шафтную съемку в пределах речного ложа.
Прежде чем приступить к полевому исследованию речных ак-
вальных комплексов, попытаемся хотя бы в виде очень беглых
зарисовок дать «портреты» главных представителей речных гидро-
фитов — водных растений.
Зона прибрежных растений — заросли низкой поймы. Двига-
ясь по пойме к реке мы, конечно же, не сразу «сваливаемся» в
речку: сначала нам предстоит пробраться через заросли низкой
прирусловой поймы, которые часто состоят из осок (рис. 4.13).
Осока острая, осока вздутая и осока черная — очень типичные
краевые виды, с удовольствием поедаемые и водяной крысой, и
кряквой.
Хвощи топяной и приречной — два других хорошо узнаваемых
растения; нередко образуют чистые ряды, окаймляющие глини-
стые и топкие иловатые побережья малых рек.
Красив и легко определяем тростник обыкновенный, создаю-
щий настоящие джунгли в рост человека и выше. Этот многолет-
ний злак украшает речку все лето, превращаясь осенью в готовый
сухоцвет для зимнего букета с длинным ломким стеблем и раски-
дистым колосом соцветья.
Манник большой (наплывающий, водяной) — развивается на мел-
ководьях в затонских частях перекатов и узнается по характерно-
му сиреневому оттенку соцветий (рис. 4.14).
Рогоз узколистный (широколистный) — всем известный «фила-
тик» (часто по ошибке именуемый в народе камышом), страдает
из-за роскошных бархатистых «султанчиков», которые простоят в
вазе всю зиму, — также прибрежное растение низкой поймы.
Аир пахучий — переселенец из юго-восточной Азии, хорошо
чувствует себя на отмелях сибирских речек; высокое растение:
140
«типажные» листья окружают округлый цветоносный стебель, в
белым мелким цветкам, собранным в щитковидную
141
узнается
по метелку.

Калужница болотная — с почковидными листьями и много-
численным желтыми цветками «тянет» к сырым лугам низкой
поймы, иногда добираясь до русла.
В стороне от течения — зона тихих речных стариц и затонов
(рис. 4.15).
Белокрыльник болотный — растение с округлым «кожистым»,
хорошо «прожилкованным» листом (по форме — опрокинутое
сердечко), почти вертикально посаженным на короткий стебель;
образует целые заросли на «стареющих водоемах» — отшнурован-
ных старицах, заиленных протоках, в затонах рек. К середине лета
142
формирует красивую ярко-красную «шишечку» на фоне белого
«манжета».
Вахта трехлистная — многолетняя трава с простертыми вер-
тикально стеблями, несущими только очередные тройчатые ли-
стья и бледно-розовые «звездочки» пятичленных цветков.
143

Час/пуха подорожниковая — вполне оправдывает свое название
и с первого взгляда действительно напоминает «подорожник»,
взбежавший на пляжевую отмель, однако собранные в сложный
зонтик мелкие белые цветки на высоком стебле-цветоносе не по-
зволят вам ошибиться.
Жеруха земноводная — красивая трава с лировидно рассечен-
ными надрезными верхними листьями, цельными или перисто-
надрезными, с золотисто желтыми цветами.
Вех ядовитый — растение, которое можно встретить и на низ-
кой пойме, и на бечеве, и в русле на сплавинах. Корневище тол-
стое, белое, разделенное на перегородки, сильно ядовито (это та
самая цикута, которой, по легенде, отравился Сократ). Мелкие
цветочки собраны шарами в белый сложный зонтик, стебель труб-
чатый полый.
Сабельник болотный — может быть встречен с одинаковым ус-
пехом и по берегам рек, и на болоте. Длинные ползучие побеги
несут пятипалые сложные листья; на конце — правильный цветок
с винно-красными лепестками, выдающий принадлежность к ро-
зоцветным.
Стрелолист обыкновенный — полупогруженный чрезвычайно
пластичный вид, может селиться как в тихих затонах, так и в
плесовых лощинах, при этом морфология растения меняется: ха-
рактерные крупные листья-стрелы заменяются длинными листь-
ями «лентами». Соцветие головчатое.
Ежеголовник ветвистый — часто образует вместе со стрелоли-
стом дружные густые заросли. Желтые цветки собраны в шаровид-
ные головки, поочередно располагающиеся на стебле-цветоносе.
Листья плотные, нередко трехгранные, линейные.
Телорез алоэвидный — может селиться на мелководье тихих за-
тонов, в заиленных отшнуровавшихся протоках и старицах, фор-
мируя очень живописные заросли. Наилучшего развития группи-
ровки телореза достигают после ряда маловодных лет с коротки-
ми и низкими паводками, чем обусловлено усиленное накопле-
ние наилка в руслах.
Водяной орех — растение с гибким, плавающим в воде стеблем,
который крепится ко дну, как якорем, прошлогодним орехом. При
подъеме уровня воды водяной орех может отрываться от грунта и
превращаться в свободно плавающее растение; влекомый потоком,
он достигает необходимой глубины и укореняется в спокойной зоне.
Русловая зона погруженных растений — плесовые лощины,
корыта перекатов, подводные части отмелей (рис. 4.16).
Эта зона обычно захватывается представителями обширного
семейства рдестовых, каждый из которых в зависимости от конк-
ретных условий может образовывать более или менее обширные
заросли. Цветы у всех рдестовых собраны в мутовчатых кистях или
зонтиковидных соцветиях. Легче всего рдесты различаются по форме
144
листовой пластинки и ее «посадке» на побег. Часто в группиров-
ках рдестов представлено сразу несколько видов, образующих гу-
стые подводные луга с разнообразными формами жизни. Рыба
откладывает здесь икру, а вышедшая из нее молодь не торопится
покидать дремучие заросли, служащие для нее надежным укры-
тием. Сюда же в поисках обильного корма заходят и взрослые рыбы.
Рдест пронзеннолистный — один из наиболее широко распрост-
раненных рдестов, также занимает средние по глубине (1,5 — 2,0 м)
плесовые лощины на малых реках, хотя его можно встретить и на
мелководьях. Название получил за своеобразную форму листовой
145