ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.05.2019
Просмотров: 22720
Скачиваний: 687
десятки или сотни раз) превышала площадь единовременно
использовавшихся подсек. Даже при небольшой плотности
населения значительная доля удобных для земледелия земель
в южной и средней тайге была так или
иначе вовлечена в подсечно-огневое земледелие. Тем не менее
основная нагрузка приходилась именно на леса, удобные для зем-
леделия (в том числе и с точки зрения доступности).
Характер лесов, формирующихся под воздействием часто и в
течение длительного времени повторяющихся пожаров и едини-
чных пожаров, разделенных многими веками естественной (окон-
ной) динамики, существенно различается (рис. 8.2). Системати-
ческое, продолжающееся в течение многих столетий воздействие
пожаров приводит к формированию светлохвойных таежных ле-
сов, в лесах которых преобладают «пирогенные» породы: сосна
(повсеместно) и лиственница (на северо-востоке европейской
России). Для сосны и лиственницы свежие гари или участки леса,
пройденные низовыми пожарами, представляют оптимальные
условия для возобновления и развития молодых поколений. Взро-
слые же деревья сосны и лиственницы способны переживать даже
довольно сильные низовые пожары.

Однако семена сосны и лиственницы относительно тяжелы и
распространяются ветром сравнительно на небольшие расстоя-
ния (в лучшем случае на сотни метров). В результате в условиях,
когда пожары случаются крайне редко, а участки новых и старых
гарей разнесены в пространстве на многие километры, господ-
ство на участках гарей получают другие пионерные породы, семе-
на которых могут преодолевать значительные расстояния или ко-
торые входят в состав естественных лесов и в условиях отсутствия
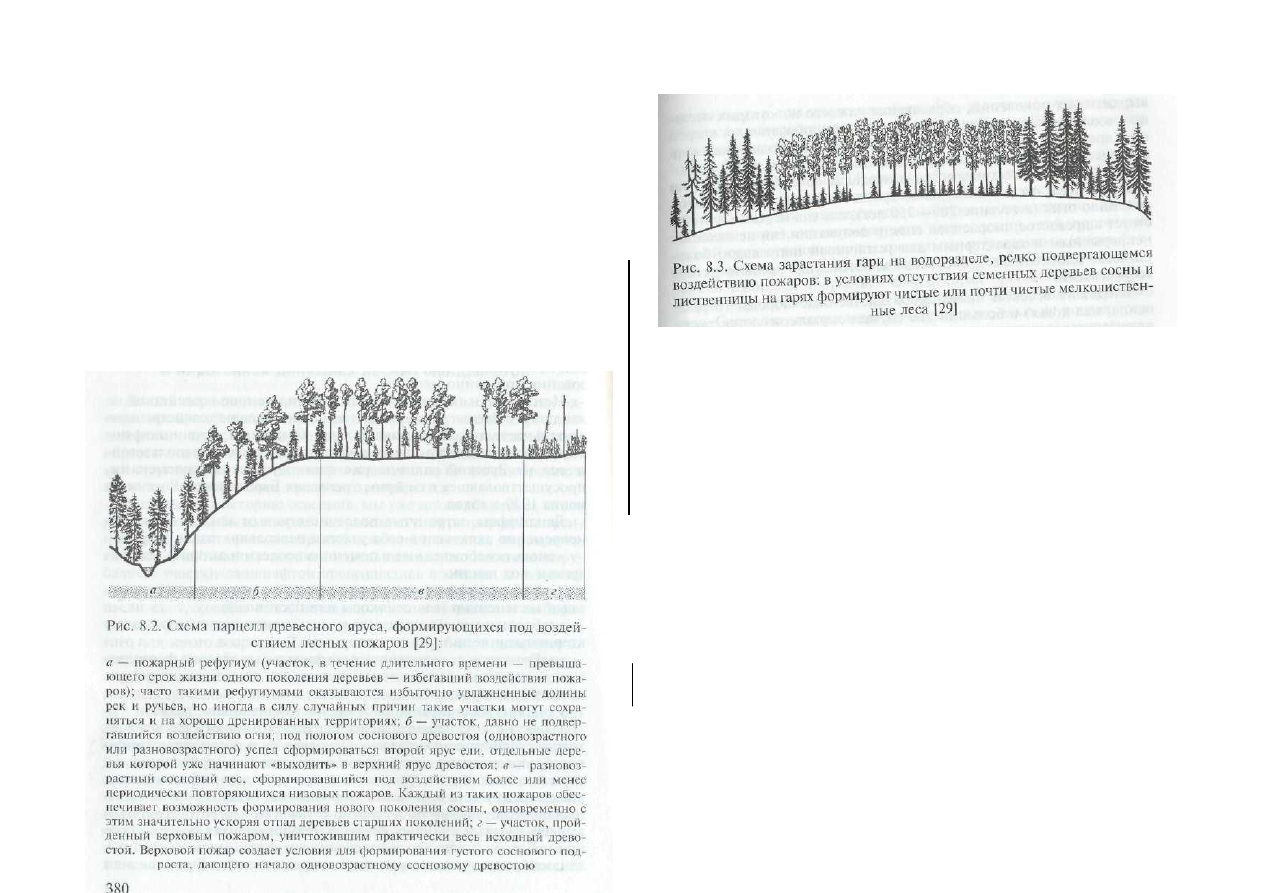
пожаров: береза и осина. Поэтому для обширных водораздельных
пространств, где в прошлом воздействие пожаров было не столь
частым, а большинство источников семян сосны и лиственницы
(там, где они были) вырублено, в течение последних полутора
столетий характерно зарастание гарей березой и осиной (рис. 8.3).
Таким образом, под влиянием пирогенного фактора трансфор-
мации лесные ландшафты превращаются в сложную мозаику с
разной частотой выгорающих участков, в которую, как правило,
вкраплены пожарные рефугиумы — участки, в силу разных причин
избежавшие пожара в течение последних нескольких столетий и
характеризующиеся, как правило, оконной (вывальной) струк-
турой древостоя. Поскольку воздействие огня определяет все ос-
новные особенности структуры и динамики конкретного участка
леса (возрастной состав древостоя, интенсивность гибели деревь-
ев, количество валежа и мертвой органики на поверхности по-
чвы, состав и структуру нижних ярусов леса), постольку участки,
испытавшие пожары разных типов (верховые, низовые) и интен-
сивности (сильные, слабые), различаются по этим параметрам и
могут формироваться как одновозрастные, простые по структуре
урочища, так и разновозрастные, состоящие из нескольких от-
дельных поколений (29].
При этом на наиболее часто выгорающих участках
формируются сосновые леса (на северо-востоке Русской
равнины
—
лиственничные),
характеризующиеся
преобладанием нескольких четко


выраженных поколений, образование каждого из которых связа-
но с воздействием низового пожара. Для ели, образующей второй
ярус древостоя на участках, длительное время не подвергавшихся
воздействию огня, характерно преобладание группы близких по-
колений с разбросом возрастов в пределах 60 — 80 лет. Даже на
участках леса, весьма длительное время не подвергавшихся воз-
действию огня (в течение 200 — 250 лет), где ель нередко господ-
ствует в древостое, возрастной спектр популяции ели не является
непрерывным и характерным для устойчивой популяции; боль-
шинство таких еловых древостоев являются лишь условно-разно-
возрастными.
Входящие в состав таких лесов лиственные породы (береза,
осина, ива козья) в большинстве случаев характеризуются также
прерывистыми возрастными спектрами с наличием одного или
нескольких четко выраженных поколений.
В лесных ландшафтах с пирогенной динамикой специфический
почвенный микрорельеф, образующийся при вываливании дере-
вьев с корнями, развит слабо, что связано с преобладанием усыха-
ния деревьев на корню над вываливанием живых старых деревьев.
Для них также характерно отсутствие мощных органогенных по-
чвенных горизонтов на незаболоченных участках и относительно
малое количество мертвого органического вещества на поверхно-
сти почвы на участках, не относящихся к пожарным рефугиумам.
Поэтому влагоудерживающая способность этих лесов существенно
меньше, чем у лесов с четко выраженной оконной динамикой.
Крайне неравномерный процесс гибели старых деревьев на
различных участках леса регулируется здесь сразу несколькими
факторами: воздействием пожаров (в том числе и низовых палов,
вызывающих усыхание ослабленных и наиболее поврежденных
огнем деревьев), гибелью старых и ослабленных деревьев от вре-
дителей и болезней, воздействием экстремальных погодных усло-
вий (засух, особенно актуальных для участков с маломощными и
бедными органическим веществом почвами).
Под воздействием современной хозяйственной деятельности
леса с пирогенной динамикой существенно изменяются. Пожары,
возникающие вокруг лесосек и дорог, коренным образом изме-
няют веками складывавшуюся структуру таежных ландшафтов:
изменяется частота пожаров, исчезают пожарные рефугиумы,
упрощается характер лесной мозаики.
Веками складывавшееся равновесие нарушается катастрофи-
ческим образом, и предсказать, к каким последствиям для био-
логического и ландшафтного разнообразия таежных лесов приве-
дут такие последствия хозяйственной деятельности человека, не
всегда представляется возможным.
Как мы уже могли убедиться, сосна обладает ярко выраженной
устойчивостью к пожарам, ель, напротив, погибает сразу же по-
еле пожара, в том числе низового. Но, в отличие от сосны, ель
обладает высокой теневыносливостью, она способна вырасти под
древесным пологом самой высокой сомкнутости.
Эти биологические особенности двух основных лесообразующих
пород центра и севера ЕТР вполне объясняют последовательность
и характер сукцессии, определяющих динамику лесных ландшаф-
тов. Сосна первой осваивает все незанятые местоположения (в осо-
бенности гари), но возможности закрепления этой породы на
долгий срок ограничены экстремальными условиями (очень бед-
ных экотопов). Под пологом сосны, а также сопутствующей ей
березы начинает подрастать ель, которая постепенно осваивает
второй ярус древостоя, а затем проникает и в первый (верхний)
ярус. Сосна не выдерживает конкуренции с елью на почвах сред-
ней и повышенной трофности, поэтому подросту сосны здесь уже
не вырасти (если только не пойдет очередной пожар). Такой ре-
жим существования сосняков назван пирогенной стабильностью [29].
О связи большинства типов сосняков с периодическими пожара-
ми свидетельствует почти полное отсутствие данных о развитии
таежных сосновых лесов ЕТР до стадии абсолютно разновозраст-
ного древостоя.
В свете изложенного выше вопрос о коренных лесах ЕТР транс-
формируется в вопрос о том, какого возраста леса следует считать
коренными, т.е. сформировавшимися без влияния человека, если
первые следы подсечного земледелия относятся к началу нашей
эры? С точки зрения естественной динамики ландшафта корен-
ными следует считать леса, существование которых определяется
господствующей тенденцией лесовосстановительных процессов —
фитоценотических смен. В этом случае ельники следует признать
коренными сообществами для большинства типов ландшафтов
региона.
Таким образом, подавляющее большинство лесов таежной зоны
Европейской России характеризуется сильной преобразованно-
стью в результате хозяйственной деятельности человека, они дол-
жны быть отнесены к вторичным лесам. Точную долю вторичных
лесов в составе таежного лесного покрова оценить трудно, в пер-
вую очередь, из-за отсутствия общепризнанных критериев отне-
сения лесов к вторичным или первичным (коренным). Так, на-
пример, нет единого представления о том, до какой стадии вос-
становления естественной структуры леса после нарушения этот
лес относится к вторичным лесам или в какой степени преобра-
зованный выборочными рубками лес может считаться вторичным.
Однако определенные категории лесов однозначно понимаются
как вторичные практически во всех исследованиях (например, леса,
образованные первым поколением деревьев на месте вырубок, заб-
рошенных сельскохозяйственных угодий, карьеров и т.д.). Даже
при самом «узком» определении вторичных лесов можно утверж-
383

дать, что вторичные леса в таежной зоне образуют общий фон, а
первичные {коренные, старовозрастные) леса — вкрапления или от-
дельные массивы внутри этого фона.
Длительность сукцессии елового леса в южной тайги оценива-
ется в 120—150 лет, причем этот период одинаков при восстанов-
лении леса как после рубки, так и после пожара. По-видимому,
указанные сроки можно считать характерным временем форми-
рования темнохвойного таежного ельника со всеми присущими
ему признаками: видовым составом, строением почвенного про-
филя, микрорельефом, мезоклиматом и т.п. Если же считать в
качестве финальной стадии разновозрастный ельник, то для его
формирования требуется не менее 300 лет [8], что практически
исключено при современном характере освоения Европейской
территории России. Следовательно, длительное естественное раз-
витие лесов в регионе является скорее исключением, а не правилом, и
наблюдаемое сегодня разнообразие лесных сообществ поддерживается
за счет периодических и разновременных внешних воздействий, как
правило, рубок различного вида. Ход следующих за ними лесовосста-
новительных сукцессии во многом определяется соседством
срубленных (сгоревших) участков леса с сохранившимися участ-
ками — источниками семян той или иной древесной породы. Так,
ель не будет восстанавливаться даже в подходящих для нее ланд-
шафтных условиях, если ельники в округе полностью отсутствуют
в радиусе десятка километров.
В настоящее время практически во всей лесной зоне россий-
ского центра и севера преобладают березовые и осиновые леса,
представляющие собой антропогенные варианты естественных
сукцессии. При сведении хвойных пород осина и береза обычно
обгоняют самосев других пород и со временем (через 25 — 30 лет)
выходят в господствующий ярус. Осина благодаря корнеотпры-
сковому образованию побегов получает особенное преимущество.
Кроме того, береза и осина — деревья быстрорастущие, светолю-
бивые; они обладают огромной семенной продуктивностью, во
много раз превосходящей продуктивность ели. Мелкие крылатые
плодики березы и крохотные семена осины, снабженные длин-
ными волосками, легко распространяются ветром на значитель-
ные расстояния.
Береза — порода, не требовательная к богатству почвы и уме-
ренно требовательная к влаге, но избегающая очень сухих место-
обитаний. Осина развивается лучше на достаточно увлажненных и
более богатых почвах. Благодаря этим экологическим особенно-
стям береза и осина повсюду первыми появляются на открытых
площадях, заселяют вырубки, гари, залежи и поляны. В насто-
ящее время урочища мелколиственных лесов распространены в
центре и на севере России по сравнению с другими лесами более
широко. Наиболее характерны леса с абсолютным господством
384
мелколиственных пород для крупных массивов концентрирован-
ных вырубок 1950-х — 1980-х годов, где не только были макси-
мально изменены условия жизни деревьев (микроклимат, почвен-
ный покров и т.д.), но часто и вырублены практически все исто-
чники семян хвойных пород деревьев.
При всем разнообразии структуры вторичных лесов, связан-
ном не только с разнообразием типов местообитаний, но и с
разнообразием видов хозяйственных воздействий, можно выде-
лить несколько основных характерных особенностей, отличающих
такие леса от естественных лесных экосистем таежной зоны. Прежде
всего, сильные антропогенные нарушения, приводящие к гибели
всего древостоя или существенной его части, приводят к суще-
ственному упрощению пространственной структуры древостоя (и,
как следствие, всех остальных ярусов леса). На наиболее нарушен-
ных хозяйственным воздействием участках, прежде всего забро-
шенных сельскохозяйственных угодьях или сплошных вырубках,
формируются наиболее простые по структуре леса с максимально
выровненным древесным пологом из мелколиственных пород,
обычно березы или серой ольхи. Любые сохранившиеся при на-
рушении фрагменты старых лесных экосистем (куртины подро-
ста, тонкомерные деревья при сплошных рубках, случайно вы-
жившие участки древостоя или деревья при пожарах) приводят к
некоторому усложнению структуры и состава формирующегося
древостоя. Выровненность и относительная одновозрастность дре-
востоя часто сохраняется и в первом поколении «коренных» дре-
весных пород, поселившемся под пологом пионерных мелколи-
ственных (например, ели, замещающей распадающийся полог мел-
колиственных пород на месте зарастающих сельскохозяйственных
угодий).
Интенсивные выборочные или условно-сплошные рубки, при
которых выбираются все крупномерные деревья, а сохраняется в
основном подрост (пусть и разновозрастный), в значительной мере
также приводят к формированию упрощенной пространственной
структуры древостоя (рис. 8.4). В таких лесах основные элементы
пространственной неоднородности древостоя и других ярусов леса
в пределах одного местообитания связаны не с проявлениями
оконной динамики, а с техногенной структурой нарушения: во-
локами, погрузочными площадками, дорогами, скотопрогонами
и т.д. [29].
Упрощенная и однообразная структура древостоя во втори-
чных лесах приводит к существенным изменениям и других яру-
сов леса. Исчезают многие виды микроместообитаний под поло-
гом леса: валеж (по крайней мере, крупный), бугры и запади-
ны, образующиеся при вываливании деревьев с корневыми си-
стемами; упрощается мозаика окон и световых пятен под поло-
гом леса. Одновременно снижается и разнообразие экологичес-
385

ких условий под пологом леса, что приводит к обеднению фло-
ристического состава, постепенному исчезновению отдельных ви-
дов растений. Точно так же восстановление исходной структуры
травяно-кустарничкового покрова происходит с существенной
задержкой по сравнению с восстановлением исходной структу-
ры древесного яруса. Это позволяет утверждать, что многие леса,
в которых уже произошло восстановление «коренной» структу-
ры древостоя после сильных антропогенных нарушений, по ха-
рактеру некоторых других компонентов леса (например, травя-
но-кустарничкового покрова) могут быть аналогичны типичным
вторичным лесам. Таким образом, учет только характера древе-
сного яруса не позволяет однозначно оценить степень нарушен-
ное™ леса хозяйственной деятельностью человека или степень
восстановления естественной структурно-динамической органи-
зации лесных экосистем [28].
Вообще смена видового состава лесных сообществ при после-
рубочных и послепожарных сукцессиях обнаруживает еще одну
характерную закономерность, которую предложено называть пра-
вилом сдвига: любое сообщество лиственного леса, развивающе-
еся после рубки хвойного древостоя, благодаря обогащению вер-
хних горизонтов почвы будет обладать более богатым составом
травяно-кустарничкового яруса по сравнению с исходным («ко-
ренным») сообществом. По мере взросления лиственного леса
такое смещение будет увеличиваться, но только до того преде-
ла, пока естественное возобновление ели не прекратит этот про-
цесс [16].
Многие антропогенные воздействия ведут в обогащению вер-
хнего горизонта лесных почв (выпас, подсека), что в сочетании
с общим осветлением определяет широкое распространение тра-
вянистых растений под пологом «бывшей» тайги: вейника лес-
ного, ландыша, костяники, перелески, звездчатки, майника и
др. Отсюда преобладание травяных инвариантов коренных лес-
ных сообществ (сосняков, ельников) над моховыми и кустар-
ничковыми. Именно такие леса, как правило, получают распро-
странение в зоне экстенсивной промысловой нагрузки вокруг
сельских населенных пунктов. В травянистых фитоценозах возоб-
новление ели затруднено и лишь по прошествии длительного
времени после смыкания полога подросших елей ель начинает
проявлять себя как сильный эдификатор — виолент, жестко ог-
раничивая (порой до четырех-пяти) число видов в напочвенном
покрове. Однако после достижения перестойного возраста (свы-
ше 150 лет) в связи с увеличением вывала в ельниках образуются
многочисленные окна, где вновь поселяются виды, требова-
тельные к свету.
Проникающие под прозрачный полог осины и березы ценные
породы постоянно вырубаются, в результате общая эволюция ле-
сов идет в крайне неблагоприятном направлении. Очевидно по-
этому еще существующие остатки хвойно-широколиственных ле-
сов должны стать предметом особой охраны, тем более что в по-
следние годы климатические условия на территории центра и се-
387