ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 27.11.2019
Просмотров: 4033
Скачиваний: 98
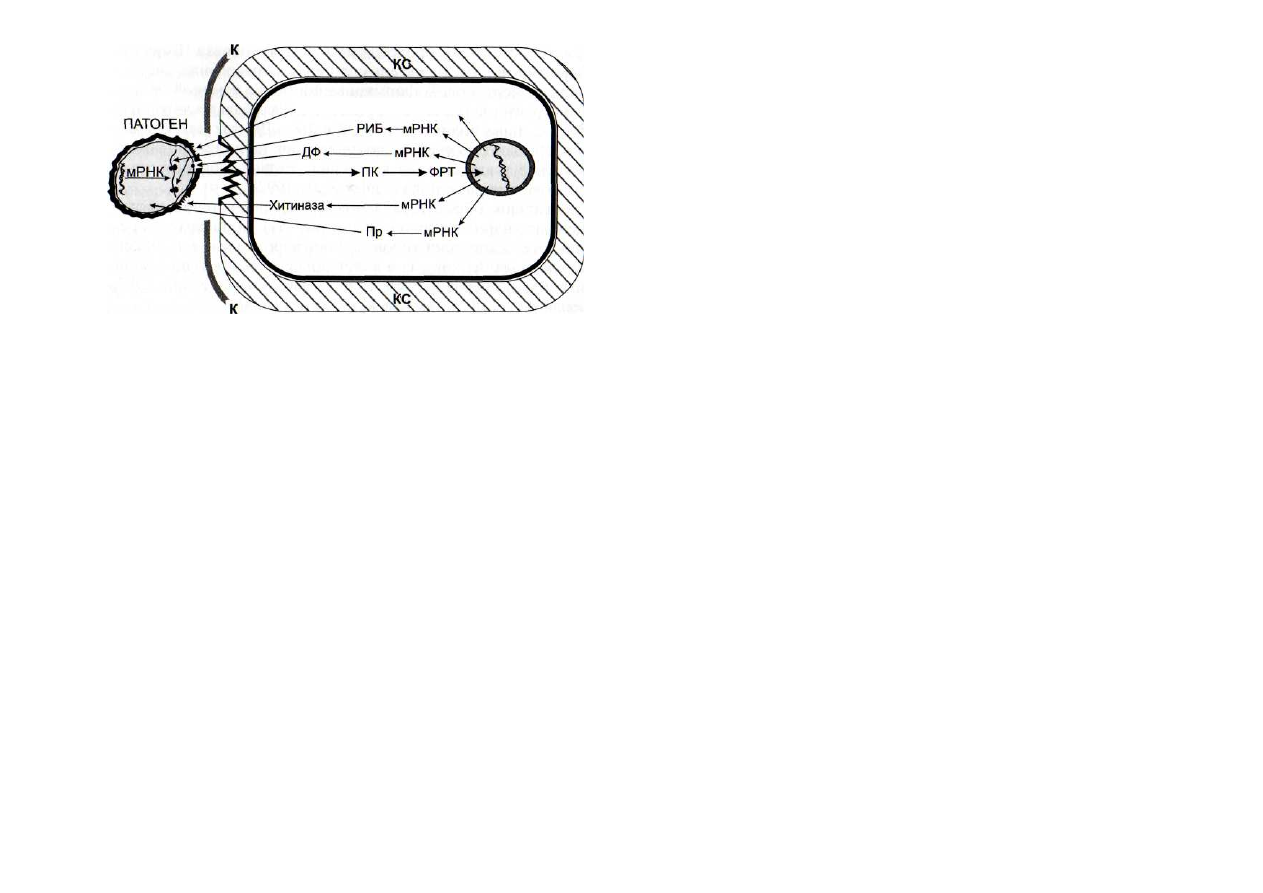
Рис. 54. Схема элиситориндуцируемого образования клетками
растений белков прямого антипатогенного действия
ДФ - дефенсины; К - кутикула; КС - клеточные стенки; ПК - про-
теинкиназы; Пр - протеиназы; РИБ - рибосомоинактивирующие бел-
ки; ФРТ - факторы регуляции транскрипции
каназ не только разрушать клеточные стенки грибов, но и
продолжать деградацию освобождающихся фрагментов хи-
тина и (3-1,3-глюканов, снижая их элиситорные способности
[Salzer et al., 1997].
Патогениндуцируемые белки растений, нарушающие
функционирование клеточной мембраны патогенов.
Под
влиянием инфицирования и ряда других неблагоприятных
факторов в растениях быстро образуются модификаторы
свойств клеточных мембран патогенных грибов и бактерий
(см. рис. 54) - относительно небольшие (от 2 до 9 кДа) по-
липептиды, подразделяемые на целый ряд семейств: тиони-
ны, дефенсины, липидпереносящие белки, хевеины, ноти-
ны, снейкины и др. [Garcia-Olmedo et al., 1998]. Список бак-
терицидных и фунгицидных полипептидов продолжает по-
полняться. Все они имеют идентичный план строения - нес-
колько дисульфидных мостиков, гидрофобное ядро, одну
протяженную а-спираль и три или четыре антипараллель-
но расположенных небольших (3-полос [Song et al., 1997;
Garcia-Olmedo et al., 1998; Fant et al.,
1999]. Так, у одного из
тионинов - у-1-пуротионина [Bruix et al., 1995], насчитыва-
ется четыре дисульфидных мостика, а-спираль, включаю-
щая участок полипептида от 16 до 28 аминокислоты, три Р-
полосы, включающие 1-6-, 31-34- и 39^7-остатки амино-
кислот. У вискотоксина дисульфидные мостики соединяют
3 и 40, 4 и 32, 16 и 26 остатки цистеина [Orru et al., 1997], а у
у-тионина из сорго - 3 и 47, 14 и 34, 20 и 41, 24 и 43 [Nitti
et al., 1995].
Примечателен факт принципиального сходства строе-
ния и относительно высокой гомологии отдельных участ-
ков этих полипептидов с нейротоксинами скорпиона и де-
фенсинами насекомых, что свидетельствует об эволюцион-
ной стабильности этих важных защитных соединений [Zinn-
Justin et al., 1996; Thevissen et al., 1997; Kushmerick et al.,
1998]. Следует ожидать, что взаимодействующие с ними ре-
цепторы клеточной мембраны бактерий и грибов также об-
ладают консервативной структурой.
Образование дефенсинов растений индуцируется не
только патогенами, но и промежуточными продуктами
сигнальных систем клеток и некоторыми стрессовыми
фитогормонами, например МеЖК и этиленом [Terras
et al., 1998; Shah et al.,
1999], однако регуляция этими со-
единениями экспрессии генов различных дефенсинов мо-
жет сильно отличаться. Так, МеЖК не действовал на син-
тез некоторых изоформ дефенсинов [Epple et al., 1997].
Как правило, салициловая кислота также не индуцирова-
ла образования дефенсинов [Epple et al., 1997; Terras et al.,
1998; Shah et aL, 1999].
При выяснении причин ингибирующего действия на
грибы тионинов и дефенсинов было обнаружено, что они
вызывают изменение мембранного потенциала клеточной
мембраны гриба [Froy, Gurevitz, 1998], усиливают погло-
щение Са
2+
, выход К
+
, подщелачивание среды [Thevissen et
al., 1996; De Samblanx et al.,
1997], ингибируют №
+
-
кана-лы
[Kushmerick et al.,
1998]. Дефенсины, тионины и липид-
переносящие белки в разной степени вызывали аггрега-
цию и усиление проницаемости для различных веществ ис-
кусственных фосфолипидных липосом [Caaveiro et al.,
1997]. Считается, что тионины могут подавлять рост гри-
бов, непосредственно (неспецифически) действуя на их
клеточные мембраны, а дефенсины - связываясь с распо-
(3-1,3-
Глюканаза-» - мРНК

ложенными в них специфическими рецепторами [Thevissen
et al., 1996].
Интересно, что грибы в ответ на действие дефенсинов
включают пока еще неизвестный механизм подавления их
образования у растений [Sharma, Lonneborg, 1996]. Этот фе-
номен проявляется через сутки и более после начала дейст-
вия антигрибных полипептидов на грибы и является еще од-
ним подтверждением гипотезы генетического пинг-понга
между патогеном и хозяином.
Патогениндуцируемые белки растений, вызывающие
нарушение процессов трансляции у патогенов.
Рибосомо-
инактивирующие белки (РИБ) относятся к широко распро-
страненным [Gasperi-Campani et al., 1985; Stirpe, Barbieri,
1986; Barbieri et al., 1993; Citores et al.,
1993; и др.] защитным
антибиотическим стрессовым белкам, синтез большинства
которых начинается после воздействия на растения биоген-
ных и абиогенных стрессоров [Stirpe et al., 1996; Rippmann
et al.,
1997; и др.] (см. рис. 54). Следует отметить, что неко-
торые из РИБ синтезируются конститутивно, например в
семенах и плодах многих растений [Vigers et al., 1991], где
вместе с другими белками (хитиназами, [3-1,3-глюканазами,
ингибиторами протеиназ) обеспечивают защиту от бакте-
рий, грибов и вирусов. В последние годы РИБ привлекли к
себе особое внимание, так как обнаружилось, что они обла-
дают противоопухолевой активностью [Langer et al., 1999;
Sharma et al., 1999].
В настоящее время известны десятки патогениндуциру-
емых представителей РИБ у растений различных семейств.
Названия этих РИБ обычно отражают родовую или видо-
вую принадлежность растений: аспарины из Asparagus offi-
cinalis [Bolognesi et al.,
1990], волкенсин из Avenia volkensii
[Sparapani et al.,
1997], бриодины из Bryonia dioica [Bolognesi
et al.,
1990], колоцины из Citrullus colocynthis [Bolognesi et al.,
1990], диантин из Dianthus caryophyllus [Hong et al., 1996],
гелонин из Gelonium multiflorum [Brigotti et al., 1999], луф-
фины и луффацилин из Luffa cylindrica [Brigotti et al., 1995],
лихнин из Lychnis chalcedonica [Bolognesi et al., 1990; Brigotti
et al.,
1995], мапалмин из Manihot palmata [Bolognesi
et al.,1990], MOR
и MOR 1 из Marah oreganus [Bolognesi et al.,
1
996], момордины [Bolognesi et al., 1996], моморхарины
| Mock et al., 1996; Wang, Ng,
1998] и моморкохин [Bolognesi
ct al.,
1990] из Momordica charantia, фитолакцин из
Phytolacca americana [Barbieri et al.,
1992], рицин из Ricinus
communis [Wang, Ng, 1998; Sharma et al.,
1999], эбулитины
из Sambucus ebulus [De Benito et al., 1995], нигритины и ниг-
рины из Sambucus nigra [Battelli et al., 1997], сапорины из
Saponaria officinalis [Bolognesi et al.,
1996], кириловины и
трихокирин из Trichosanthes kirilowii [Brigotti et al., 1995],
тритины из Triticum aestivum [Brigotti et al., 1995], синамо-
мин и камфорин из Cinnamomum camphora [Li, Chory, 1997]
и т.д.
Определение первичной структуры многих РИБ показа-
ло, что они обладают более или менее хорошо выраженной
гомологией [Funatsu et al., 1991; Wang, Ng, 1998] и что все
они могут быть подразделены на два основных типа: одно-
цепочечные (РИБ I) и двухцепочечные (РИБ II). Некото-
рые из них гликозилированы [Di Maro et al., 1999]. Молеку-
лярные массы большинства РИБ находятся в пределах
28-
32 кДа. Защита против патогенных бактерий и грибов
обеспечивается ингибирующим действием РИБ на процесс
трансляции в рибосомах, а именно блокированием фактора
элонгации [Citores et al., 1993]. Специальные исследования
молекулярного механизма подавления трансляции позволи-
ли установить, что РИБ вызывают расщепление N-связи
между рибозой и аденином, причем в специфическом нук-
леотиде А-4256, который находится в петле 28S в рибосо-
мальной РНК, входящей в состав 60S субъединицы рибосо-
мы [Fong et al., 1991; Brigotti et al., 1999]. Считается, что де-
иуринизация нуклеотида А-4256 нарушает динамическую
гибкость структуры рибосом, которая необходима для осу-
ществления синтеза очередной пептидной связи [Holmberg,
Nygard, 1996].
Накапливается все больше фактов не только о N-глико-
шдазной, но также о суперспиральзависимой эндонуклеаз-
ной [Liu, Pu, 1999], РНКазной [Obrig et al., 1985; Mock et al.,
1996], ДНКазной [Nicolas et al., 1998] активностях РИБ. Об-
наружен также новый фермент - сайт-специфическая
рРНК-лиаза, которая способна расщеплять молекулу РНК
на 3'-участке апуринового сайта [Ogasawara et al., 1999] и ра-
ботающая в комплексе с N-гликозидазой, обеспечивая не
только точечное видоизменение, но и последующее раз-
рушение рибосомной РНК патогенов. У одного из РИБ -
камфорина, обнаружена супероксиддисмутазная актив-
ность [Li, Chory, 1997].
Получены любопытные данные о связи структуры не-
которых РИБ с другими стрессиндуцируемыми белками.
Так, N-концевой участок одного из РИБ типа I отличался от
аналогичного участка хитиназы лишь одной аминокисло-
той [Di Maro et al., 1999]. Обнаружена гомология между N-
концом жасмонатиндуцируемого белка 60 кДа и каталити-
ческим доменом одного из РИБ [Fong et al., 1991]. Оказа-
лось, что последний может образовываться из белка 60 кДа
в ходе двухступенчатого процессинга.
Многие РИБ вызывают гибель не только грибов и бак-
терий, но и клеток растений и животных [Chaudhry et al.,
1994; Bolognesi et al.,
1996]. В клеточной мембране найден
специфический РИБ I-связывающий белок, причем РИБ I
не проникает через плазмалемму в неинфицированные про-
топласты, в отличие от инфицированных вирусами, что
предотвращает их размножение [Watanabe et al., 1997].
РИБ I проявляют гомологию с А-цепью РИБ II [Wang, Ng,
1998], обладающей каталитической активностью, а проник-
новение РИБ II в клетки (перенос А-цепи) обеспечивается с
помощью В-цепи, отвечающей за узнавание специфических
рецепторных белков клеточной мембраны [Sharma et al.,
1999]. В-цепь обладает галактозоспецифичным лектино-
вым доменом, узнающим галактозные остатки на поверх-
ности клеточной мембраны [Chaudhry et al., 1994].
Гены перечисленных в разделе десятков белков облада-
ют определенной видовой, тканевой и органоидной специ-
фичностью. Вид и интенсивность синтеза белков зависят от
природы элиситорных сигналов и времени, прошедшего
после начала их действия. Можно быть абсолютно уверен-
ным, что в одном опыте на определенном объекте исследо-
ватель не сможет обнаружить значительную часть этих
белков. В этом отношении представляется показательным
исследование пространственных и временных характе-
ристик транскриптов различных белков, образующихся в
листьях бобов в месте инокуляции патогенных бактерий и
на различном расстоянии от него [Meier et al., 1993]. В мес-
те инокуляции обнаруживались мРНК хитиназ, фенилала-
нин-аммиак-лиазы, халконсинтазы, в небольшой степени -
липоксигеназ. На расстоянии 0,5- 0,7 см от места инокуля-
ции было зарегистрировано высокое содержание мРНК ли-
поксигеназ, не было найдено мРНК халконсинтазы и лишь
в небольших количествах обнаруживались мРНК хитиназы
и фенилаланин-аммиак-лиазы. На еще большем удалении
можно было найти лишь мРНК липоксигеназ, но их содер-
жание было достаточно высоким. Естественно, что по мере
увеличения времени действия патогенов системные элиси-
горные сигналы распространяются на все большее расстоя-
ние и в удаленных от места инфекции клетках индуцируется
синтез защитных белков и фитоалексинов.
ИСПОЛЬЗОВАНИЕ ЭЛИСИТОРОВ И
ИНТЕРМЕДИАТОВ СИГНАЛЬНЫХ СИСТЕМ
КЛЕТОК ДЛЯ СОЗДАНИЯ ПРЕПАРАТОВ,
ПОВЫШАЮЩИХ УСТОЙЧИВОСТЬ РАСТЕНИЙ
К ПАТОГЕНАМ
Материалы, приведенные в предыдущем разделе, свиде-
тельствуют, что элиситоры, интермедиаты сигнальных сис-
тем и стрессовые фитогормоны индуцируют образование
большого набора защитных белков, в том числе фермен-
тов, катализирующих образование антипатогенных ве-
ществ небелковой природы. Некоторые из защитных со-
единений повышают устойчивость самих растений, другие
оказывают ингибирующее действие на развитие патогенов.
Все это приводит к снижению отрицательного действия
патогенов на продукционные процессы и урожай растений,
что не могло не обратить на себя внимания. Было предло-
жено достаточно много рекомендаций практического ис-
пользования препаратов, содержащих природные элисито-
ры, интермедиаты сигнальных систем и стрессовые фито-
гормоны или их химические аналоги. Часть этих предложе-
ний запатентована, и налажены выпуск и реализация анти-
фитопатогенных препаратов.
Из элиситоров чаще всего использовались арахидоновая
кислота и производные хитина - олигохитозаны. В качест-
ве сырья для получения арахидоновой кислоты используют
морских животных и некоторые органы теплокровных жи-
вотных. Первый препарат для повышения устойчивости
растений к патогенам на основе арахидоновой кислоты был
предложен более 20 лет тому назад [Метлицкий и др., 1978,
19826; Озерецковская, 1994]. Установлено, что после пред-
посевной обработки клубней картофеля или листьев в пе-
риод бутонизации значительно повышалась комплексная
устойчивость к фитофторозу, ранней сухой пятнистости,
ризоктониозу и парше. Прибавка урожая составляла в сред-
нем 25%. Обработка арахидоновой кислотой защищала
клубни картофеля и при хранении [Чаленко и др., 2001]. За-
щитное действие арахидоновой кислоты от фитопатогенов
было подтверждено при исследовании ее действия на тома-
ты и сахарную свеклу [Метлицкий, Озерецковская, 1985].
Установлено, что арахидоновая и эйкозапентаеновая
кислоты индуцируют не только локальную, но и системную
пролонгированную устойчивость картофеля к возбудителю
фитофтороза [Чалова и др., 1989]. Было также обнаруже-
но, что арахидоновая кислота повышает устойчивость к не-
матодам при выращивании растений в теплицах [Зиновьева
и др., 1996], особенно в сочетании с метилжасмонатом [Зи-
новьева и др., 1998].
В Российской Федерации налажен производственный
выпуск антипатогенных препаратов на основе арахидоно-
вой кислоты.
В последние годы широко испытывается действие на ус-
тойчивость растений к патогенам еще одного элиситора -
хитозана. Обнаружено, что максимальная фитофтороус-
тойчивость картофеля проявляется при использовании во-
дорастворимого хитозана с молекулярной массой 5 кДа
[Переход и др., 1997; Васюкова и др., 2000]. Хитозан повы-
шал также устойчивость к нематодам растений томатов в
условиях тепличного хозяйства [Зиновьева и др., 1999], при-
чем индуцировал не только локальную, но и системную ус-
тойчивость растений к фитофторе и нематодам [Васюкова
и др., 2001].
На основе хитозана 5 кДа создан препарат "Агрохит",
рекомендуемый для защиты картофеля от фитофтороза,
но, по-видимому, достаточно эффективный в защите и от
других патогенов.
Молекулярные механизмы и практические аспекты ис-
пользования хитозана для повышения устойчивости расте-
ний к различным патогенам обсуждались на 6-м Междуна-
родном симпозиуме по хитину и хитозану, проведенном в
Москве в 2001 г.
Практическое применение получили препараты на ос-
нове стрессовых фитогормонов и их синтетических анало-
гов. Достаточно широкое распространение получили
препараты на основе 2-хлорэтилфосфоновой кислоты (2-
ХЭФК), пролонгированно освобождающие фитогормон
этилен: амхел, этрел, кампозан, флорел, гидрел, дигидрел
[Кораблева, Платонова, 1995]. Установлено, что обработка
этими препаратами растений картофеля и клубней перед
закладкой на хранение усиливает покой клубней и их устой-
чивость к фитопатогенным микроорганизмам и улучшает
качество семенного материала [Метлицкий и др., 1982а; Ко-
раблева и др., 1989]. Положительные результаты от приме-
нения препаратов-доноров этилена были получены и при
хранении лука и моркови [Карякина и др., 1990; Кораблева,
Платонова, 1995].
Повышение устойчивости к болезнями продемонстри-
ровано при использовании препаратов брассиностероидов и
их аналогов [Кораблева, Платонова, 1995].
Начинается практическое использование еще одного
стрессового гормона - жасмоната (и его производного - ме-
тилжасмоната) [Зиновьева и др., 1998].
Давно известно, что экзогенный стрессовый фитогор-
мон салициловая кислота (являющаяся также интермедиа-
том НАДФН-оксидазной и NO-синтазной сигнальных сис-
тем) и ацетилсалицилат вызывают синтез защитных соеди-
нений и повышение устойчивости растений к патогенам.
Практическое использование нашел природный миметик
салициловой кислоты [Тарчевский и др., 1999] — янтарная
кислота.
Уже давно было отмечено, что янтарная кислота явля-
ется биологически активным соединением [Благовещен-
ский, 1968]. Нами в полевых условиях на более чем
10 тыс. га и в тепличных хозяйствах на многих сельскохозяй-
ственных культурах было показано, что предпосевная обра-
ботка семян или вегетирующих растений препаратами ян-
тарной кислоты приводит к повышению интенсивности
продукционных процессов и урожаев растений, что в значи-
тельной степени связано с их устойчивостью к болезням.
Технология получения янтарной кислоты, не содержа-
щей примесей тяжелых металлов, была разработана в Ка-
занском химико-технологическом институте в лаборатории
профессора А.Г. Лиакумовича, и было начато производство
препаратов на основе этого соединения.
Технология использования тех или иных элиситоров,
ннтермедиатов сигнальных систем и стрессовых фитогор-
монов по отдельности или в сочетаниях должна быть раз-
работана применительно к конкретным видам и сортам
растений [Озерецковская, Васюкова, 2002], в противном
случае может быть получен результат, противоположный
ожидаемому.