ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 06.07.2024
Просмотров: 134
Скачиваний: 0
СОДЕРЖАНИЕ
1.Классификация и номенклатура ферментов
4) Структура белковой молекулы
8)Биосинтез холерестерина и его биологическая роль.
10) .Роль желчных кислот, переваривание липидов
11) Характеристика гормонов поджелудочной железы
15) Характеристика витаминов(в2 и в5)
19)Ферменты биологического окисления
20)Взаимосвязь аминоуглеводов с обменом липидов
23) Факторы, влияющие на скорость ферментативных реакций
24.Синтез жиров из продуктов углеводного обмена
Углеводный обмен или метаболизм углеводов в организмах животных и человека. Метаболизм углеводов в организме человека состоит из следующих процессов:
Расщепление в пищеварительном тракте поступающих с пищей поли- и дисахаридов до моносахаридов, дальнейшее всасывание моносахаридов из кишечника в кровь.
Синтез и распад гликогена в тканях (гликогенез и гликогенолиз), прежде всего в печени.
Гликолиз — распад глюкозы. Первоначально под этим термином обозначали только анаэробное брожение, которое завершается образованием молочной кислоты (лактата) или этанола и углекислого газа. В настоящее время понятие «гликолиз» используется более широко для описания распада глюкозы, проходящего через образование глюкозо-6-фосфата, фруктозодифосфата и пирувата как в отсутствии, так и в присутствии кислорода. В последнем случае употребляется термин «аэробный гликолиз», в отличие от «анаэробного гликолиза», завершающегося образованием молочной кислоты или лактата.
Аэробный путь прямого окисления глюкозы или, как его называют, пентозофосфатный путь (пентозный цикл).
Взаимопревращение гексоз
Аэробный метаболизм пирувата. Этот процесс выходит за рамки углеводного обмена, однако может рассматриваться как завершающая его стадия: окисление продукта гликолиза — пирувата.
Глюконеогенез — образование углеводов из неуглеводных продуктов (пирувата, лактата, глицерина, аминокислот, липидов, белков и т. д.).
26)основные свойства ферментов. К ферментамприменимы три основных критерия, характерных и для неорганическихкатализаторов. В частности, они остаются неизмененными послереакции, т.е. освобождаясь, могут вновь реагировать с новымимолекуламисубстрата(хотя нельзя исключить побочных влияний условий среды наактивность фермента).Ферментыспособны оказывать действие в ничтожно малыхконцентрациях(например, однамолекулаферментареннина, содержащегося в слизистой оболочке желудка теленка, створаживает около 106 молекулказеиногенамолоказа 10 мин притемпературе37°С). Наличие либо отсутствиеферментаили любого другогокатализаторане оказывает влияния на величинуконстанты равновесияи свободной энергии (ΔG).Катализаторылишь повышают скорость, с которой система приближается ктермодинамическому равновесию, не сдвигая точкиравновесия.Химические реакциис высокойконстантой равновесияи отрицательной величиной ΔG принято называть экзергоническими.Реакциис низкойконстантой равновесияи соответственно положительной величиной ΔG (они обычно не протекают спонтанно) называются эндерго-ническими. Для начала и завершения этихреакцийнеобходим приток энергии извне. В живых системах экзергонические процессы обычно сопряжены с эндергоническимиреакциями, обеспечивая последние необходимым количеством энергии.
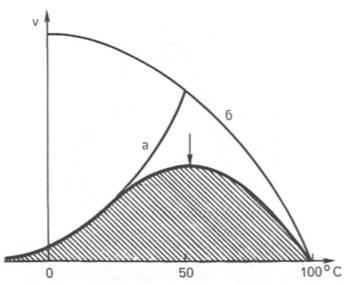
Рис. 4.16. Зависимость скорости катализируемой ферментомреакцииоттемпературы.
а - повышение скорости реакциикак функциятемпературы; б - снижениескорости реакциикак функцияденатурациибелка-фермента; стрелка указывает оптимумтемпературы.
Ферменты, являясьбелками, обладают рядом характерных для этого классаорганических соединений свойств, отличающихся от свойств неорганическихкатализаторов.
Термолабильность ферментов.Скорость химических реакцийзависит оттемпературы, поэтому катализируемыеферментамиреакциитакже чувствительны к изменениямтемпературы. Установлено, что скорость большинства биохимическихреакцийповышается в 2 раза при повышениитемпературына 10°С и, наоборот, снижается в 2 раза при понижениитемпературына 10°С. Этот показатель получил название температурного коэффициента. Однако вследствие белковойприроды ферментатепловаяденатурацияпри повышениитемпературыбудет снижать эффективнуюконцентрациюферментас соответствующим снижениемскорости реакции. Так, притемпературе, не превышающей 45–50°С,скорость реакцииувеличивается согласно теориихимической кинетики. Притемпературевыше 50°С наскорость реакциибольшое влияние начинает оказывать тепловаяденатурациябелка-фермента, приводящая к полному прекращению ферментативного процесса (рис. 4.16).
Таким образом, термолабильность, или чувствительность к повышению температуры, является одним из характерных свойствферментов, резко отличающих их от неорганическихкатализаторов. В присутствии последнихскорость реакциивозрастает экспоненциально при повышениитемпературы(см. кривую «а» на рис. 4.16). Притемпературе100°С почти всеферментыутрачивают своюактивность(исключение составляет, очевидно, только одинферментмышечной ткани– миокиназа, которая выдерживает нагревание до 100°С). Оптимальной для действия большинстваферментовтеплокровных животных являетсятемпература40°С; в этих условияхскорость реакцииоказывается максимальной вследствие увеличения кинетической энергии реагирующихмолекул. При низкихтемпературах(0°С и ниже)ферменты, как правило, не разрушаются, хотяактивностьих падает почти до нуля. Во всех случаях имеет значение время воздействия соответствующейтемпературы. В настоящее время дляпепсина,трипсинаи ряда другихферментовдоказано существование прямой зависимости
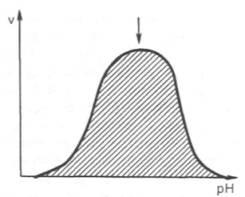
Рис. 4.17. Зависимость скорости катализируемой ферментомреакцииот рН (стрелка указывает оптимум рН).
между скоростью инактивации ферментаи степеньюденатурации белка. Следует отметить, что на термолабильностьферментовопределенное влияние оказываетконцентрациясубстрата, рН среды и другие факторы.Зависимость активности ферментовот рН среды.Ферментыобычно наиболее активны в пределах узкой зоныконцентрацииводородныхионов, соответствующей для животныхтканейв основном выработанным в процессеэволюциифизиологическим значениям рН среды 6,0–8,0. При графическом изображении на кривой колоколообразной формы имеется определенная точка, в которойферментпроявляет максимальнуюактивность; эту точку называют оптимумом рН среды для действия данногофермента(рис. 4.17). При определении зависимостиактивности ферментаотконцентрацииводородныхионовреакциюпроводят при разных значениях рН среды, обычно при оптимальнойтемпературеи наличии достаточно высоких (насыщающих)концентрацийсубстрата. В табл. 4.3 приводятся оптимальные значения рН среды для рядаферментов.
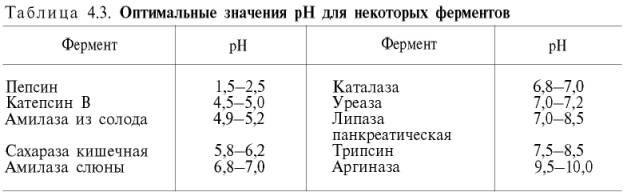
Из данных табл. 4.3 видно, что рН-оптимум действия ферментовлежит в пределах физиологических значений. Исключение составляютпепсин, рН-оптимум которого 2,0 (при рН 6,0 он не активен и не стабилен). Объясняется это, во-первых, структурной организациеймолекулыферментаи, во-вторых, тем, чтопепсинявляется компонентом желудочного сока, содержащего свободнуюсоляную кислоту, которая создает оптимальную кислую среду для действия этогофермента. С другой стороны, рН-оптимум аргиназы лежит в сильнощелочной зоне (около 10,0); такой среды нет вклеткахпечени, следовательно, in vivo аргиназа функционирует, по-видимому, не в своей оптимальной зоне рН среды.
Согласно современным представлениям, влияние изменений рН среды на молекулуферментазаключается в воздействии на состояние и степень ионизации кислотных и основных групп (в частности, СООН-группы дикар-боновыхаминокислот, SH-группыцистеина, имидазольногоазотагисти-дина, NH2-группы лизинаи др.). При резких сдвигах от оптимума рН средыферментымогут подвергаться конформационным изменениям, приводящим к потереактивностивследствиеденатурацииили изменения зарядамолекулыфермента. При разных значениях рН средыактивный центрможет находиться в частично ионизированной или неионизированной форме, что сказывается натретичной структуре белкаи соответственно на формировании активного фермент-субстратного комплекса. Имеет значение, кроме того, состояние ионизациисубстратови кофакторов.
Специфичностьферментов.Ферментыобладают высокойспецифичностьюдействия. Это свойство часто существенно отличает их от неорганическихкатализаторов. Так, мелкоизмельченныеплатинаипалладиймогут катализироватьвосстановление(с участием молекулярноговодорода) десятков тысячхимических соединенийразличной структуры. Высокаяспецифичностьферментовобусловлена, как было отмечено, конфор-мационной и электростатическойкомплементарностьюмеждумолекуламисубстратаиферментаи уникальной структурной организациейактивного центра, обеспечивающими «узнавание», высокое сродство и избирательность протекания одной какой-либореакциииз тысячи другиххимических реакций, осуществляющихся одновременно в живыхклетках.
В зависимости от механизма действия различают ферментыс относительной (или групповой) и абсолютнойспецифичностью. Так, для действия некоторых гидролитическихферментовнаибольшее значение имееттип химической связивмолекулесубстрата. Например,пепсинв одинаковой степени расщепляетбелкиживотного и растительного происхождения, несмотря на то что этибелкисущественно отличаются друг от друга как по химическому строению и аминокислотному составу, так и по физико-химическим свойствам. Однакопепсинне расщепляет ниуглеводы, нижиры. Объясняется это тем, что точкой приложения, местом действияпепсинаявляется пептидная —СО—NH-связь. Для действиялипазы, катализирующейгидролизжировнаглицеринижирные кислоты, подобным местом является сложноэфирная связь. Аналогичной групповойспецифичностьюобладаюттрипсин,химотрипсин,пептидазы,ферменты, гидроли-зующие α-гликозидные связи (но не β-гликозидные связи, имеющиеся вцеллюлозе) вполисахаридах, и др. Обычно этиферментыучаствуют впроцессе пищеварения, и их групповаяспецифичность, вероятнее всего, имеет определенный биологический смысл. Относительнойспецифичностьюнаделены также некоторые внутриклеточныеферменты, напримергексокиназа, катализирующая в присутствииАТФфосфорилиро-вание почти всехгексоз, хотя одновременно вклеткахимеются и специфические для каждойгексозыферменты, выполняющие такое же фос-форилирование (см. главу 10).
Абсолютной специфичностьюдействия называют способностьферментакатализировать превращение только единственногосубстрата. Любые изменения (модификации) в структуресубстратаделают его недоступным для действияфермента. Примерами такихферментовмогут служить аргиназа, расщепляющая в естественных условиях (ворганизме)аргинин,уреаза, катализирующая распадмочевины, и др.
Имеются экспериментальные доказательства существования так называемой стереохимическойспецифичности, обусловленной существованием оптически изомерных L- и D-форм или геометрических (цис-и транс-)изомеровхимическихвеществ. Так, известны оксидазы L- и D-аминокислот, хотя в природныхбелкахобнаружены только L-ами-нокислоты. Каждый из видов оксидаз действует только на свой специфическийстереоизомер.
![]()
![]()
Наглядным примером стереохимической специфичностиявляется бактериальная аспартатдекарбоксилаза, катализирующая отщепление СО2 только от L-аспарагиновой кислотыс превращением ее в L-аланин. Сте-реоспецифичность проявляютферменты, катализирующие и синтетическиереакции. Так, изаммиакаи α-кетоглутарата во всех живыхорганизмахсинтезируется L-изомерглутаминовой кислоты, входящей в состав природныхбелков. Если какое-либо соединение существует в форме цис-и транс-изомеров с различным расположением группатомоввокругдвойной связи, то, как правило, только один из этихгеометрических изомеровможет служить в качествесубстратадля действияфермента. Например,фумаразакатализирует превращение только фумаровойкислоты(трансизомер), но не действует намалеиновую кислоту(цис-изомер):

Таким образом, благодаря высокой специфичностидействияферментыобеспечивают протекание с большой скоростью лишь определенныххимических реакцийиз огромного разнообразия возможных превращений в микропространствеклетоки целостноморганизме, регулируя тем самым интенсивностьобмена веществ.
29)биосинтез триглицеридов. Известно, что скорость биосинтеза жирных кислот во многом определяется скоростью образования триглицеридов ифосфолипидов, так как свободные жирные кислоты присутствуют втканях и плазме крови в небольших количествах и в норме не накапливаются.
Синтез триглицеридов происходит из глицерина и жирных кислот(главным образом стеариновой, пальмитиновой и олеиновой). Путьбиосинтеза триглицеридов в тканях протекает через образование α-глице-рофосфата (глицерол-3-фосфата) как промежуточного соединения.
В почках, а также в стенке кишечника, где активность ферментаглицеролкиназы высока, глицерин фосфорилируется за счет АТФ с образованием глицерол-3-фосфата: