Файл: Научноисследовательская работа студентки 4 курса Направления подготовки 06. 03. 01 Биология Профиль Биохимия.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 04.12.2023
Просмотров: 194
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
СОДЕРЖАНИЕ
Уравнения кривой оксигенации гемоглобина
Определение параметров взаимодействия гемоглобина с протонами водорода (эффект Бора)
Определение начальных условий для распределения кислорода в гемоглобине
ГЛАВА 2 МЕХАНИЗМЫ РЕГУЛЯЦИИ СВЯЗЫВАНИЯ ГЕМОГЛОБИНА С КИСЛОРОДОМ
2.1. Модельные представления, используемые для описания взаимодействия гемоглобина с кислородом
2.2. Свободная энергия системы гемоглобин – лиганды
2.3. Описание неравновесных процессов взаимодействия гемоглобина с кислородом
надышавшийся угарным газом, задыхается, несмотря на то, что во вдыхаемом им воздухе достаточно много кислорода. Нужно срочно избавить организм от СО – вынести отравленного человека на свежий воздух, интенсивное дыхание выводит угарный газ из кровотока. Сходный эффект вызывает другая молекула – анион цианистой кислоты (CN‾). Самая знаменитая соль этой кислоты – цианистый калий. Механизм отравления цианистым калием тот же, что и угарным газом, – блокирование связи гемоглобина с кислородом.
Известно, что одна молекула гемоглобина состоит из четырех мономеров, мономер имеет тем область присоединения лиганда (молекул кислорода, углекислого газа и др.) и глобин белковую часть, взаимодействующую с протонами водорода [1]. Чем может находиться в нескольких состояниях, а именно: к гему присоединен кислород; к тему присоединен углекислый газ либо он может быть свободен. Каждый гем может обратимо присоединить одну молекулу кислорода. Соответственно молекула гемоглобина может присоединить 4 молекулы кислорода.
На рисунке 12 показаны типы взаимодействий, учитываемые в модели,
где Fe ион железа, L лиганд (молекула кислорода или углекислого газа), Н протон водорода. Стрелками указаны взаимодействия, учитываемые в модели. 1 -взаимодействие лигандов с гемом, 2 кооперативное взаимодействие молекул кислорода между собой, 3 взаимодействие протонов водорода с глобином, 4 конформационное взаимодействии макромолекулы гемоглобина с протонами водорода.
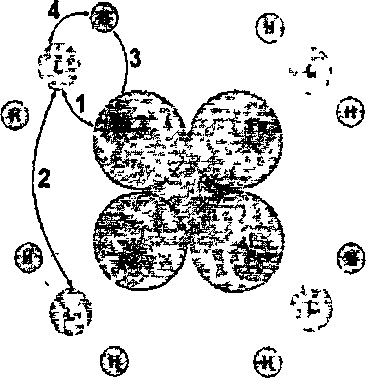
Рисунок 12 – Схематичное изображение предложенной макромолекулы гемоглобина. Цифрами обозначены учитываемые взаимодействия.
Для описания модели воспользуемся базовой моделью дефектообразования в конденсированных средах, описанной в работах С. В. Булярского [2]. Данная модель вполне подходит для описания процесса взаимодействия гемоглобина с кислородом.
В соответствии с вышесказанным примем следующую модель взаимодействия гемоглобина с кислородом.
В соответствии с вышесказанным примем следующую модель взаимодействия гемоглобина с кислородом. Обозначим все возможные состояния макромолекулы гемоглобина следующей системой индексов:
{i, j, k,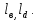 } (4)
} (4)
Численное значение индекса i соответствует количеству присоединенных молекул кислорода, j количеству молекул углекислого газа, индекс к количеству молекул присоединение еще одного лиганда.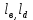
индексы зарядового состояния связи (донорной и акцепторной), способной принимать и отдавать водород, для аминокислоты.
Одна из важнейших функций белков состоит в специфическом катализе химических реакций. Лигандом в этом случае служит молекула субстрата, связывание которой ферментом необходимая предпосылка химической реакции.
Ферменты способны очень сильно ускорять химические реакции значительно сильнее, чем любые искусственные катализаторы. Столь высокую эффективность можно приписать нескольким факторам. Во-первых, ферменты увеличивают локальную концентрацию молекул субстрата в каталитическом центре и удерживают соответствующие атомы в ориентации, необходимой для последующей реакции.
Но наиболее важное значение имеет тот факт, что часть энергии связывания непосредственно используется для катализа. Дело в том. что молекулы субстрата, перед тем как превратиться в продукты реакции, проходят через ряд промежуточных форм с измененной геометрией и измененным электронным распределением.
Свободная энергия всех этих промежуточных форм и особенно наименее стабильных переходных состояний существенно снижена, если молекула связана с поверхностью фермента. Обычно ферменты имеют значительно большее сродство к нестабильным переходным состояниям субстратов, чем к их стабильным формам. Используя энергию связывания, ферменты помогают субстратам принять определенное переходное состояние и таким образом значительно ускоряют одну определенную реакцию.
Свободная энергия связывания составляет —7,1 ккал/ моль, что соответствует константе сродства 10 л/моль. Присоединение лиганда перетаскивает белок К из обычной неактивной формы К в каталитически активную форму К.
И. Линейная молекула имеет очень большое число идентичных центров связывания лиганда X. Свободная энергия взаимодействия между лигандами, связанными с расположенными по соседству друг с другом центрами, равна е.
АО принимает участие в связывании с одной определенной комбинацией орбит свободных электронных пар лигандов, что приводит к накоплению примерно 0,5 единицы электронного заряда на N1 и к сравнительно небольшой энергетической стабилизации, так как обычные энергии связей бывают, по-видимому, порядка 30 ккал. Обе эти величины малы вследствие относительно большого различия в энергии между 4 и электронами свободных пар у лигандов,
Конечно, прямой доступ к иону железа для лигандов закрыт аминокислотами, особенно дистальным гистидином. Как уже отмечалось, один из атомов азота имидазольного кольца гистидина обращен к железу, а другой фактически находится на поверхности, так что этот гетероцикл может работать как своего рода люк, перекрывающий лигандную полость.
Поэтому связывание любого лиганда представляет собой сложный процесс, включающий промежуточные изменения конформации белка, например поворот гистидина Е7 вокруг его связи Са —Сз или небольшое искажение структуры спирали Е [161]. Тем не менее скорость связывания кислорода исключительно велика. Константа скорости реакции второго порядка при 20°С для различных миоглобинов находится в интервале 1,0-10 — 1,9-10 дм -моль с [определенные к настоящему времени значения свободной энергии активации для этих процессов составили в трех случаях 23,0, 23,0 и 29,3 кДж/моль (5,5, 5,5 и 7,0 ккал/моль) соответственно], а константы скорости для изолированных, но слегка модифицированных аи 3-цепей составили 5-10 — 8-10 дм моль с , тогда как для мономерного гемоглобина hironomus получено более высокое значение 3-10 дм -моль 1-с [6]. Для гемоглобинов кинетика реакции имеет сложный характер вследствие изменений четвертичной структуры, однако константы скорости и в этом случае попадают в интервал 10 — 10 дм моль с . Константы скорости отщепления кислорода составляют 10—70 с , а соответствующие энергии активации равны 80—88 кДж/моль (19—21 ккал/моль) для миоглобинов и 10— 15 с и 67—105 кДж/моль (16—25 ккал/моль) для большинства гемоглобинов (эти значения сильно зависят от pH). Библиографию по этому вопросу см. в работе [8].
Даже если гистидин существенно уменьшает величину константы скорости, которая была в отсутствие белка, наблюдаемые скорости вполне достаточны для физиологических потребностей. Мутантные гемоглобины, в которых гистидин замещен на аргинин или тирозин, обнаруживают несколько более высокие скорости, особенно в реакциях с СО [8]. Некоторые гемоглобины с очень малыми константами скорости диссоциации ( 10 с 1), которые явно не могут функционировать как переносчики кислорода, встречаются у нематод
Вывод, что вода может вступать в реакцию через 4 /-орбиту, в то время как анионы не могут сделать этого, можно
понять на основании теории Крейга о сжатии, вызываемом во внешних -орбитах электроно-притягивающими (электроотрицательными) лигандами [19]. Анионы отталкивают электроны (электроположительны), хотя в связанном виде они становятся менее электроположительными. Внешние й -орбиты не пригодны для связывания таких лигандов из-за своей большой протяженности и низкой амплитуды, вследствие чего, несмотря на полное перекрывание, интеграл перекрывания с относительно маленькими и подразделенными орбитами лигандов очень мал. Однако молекула воды, почти нейтральная в свободном состоянии, становится заряженной положительно и потому электронопритягивающей (электроотрицательной), когда она связана. Такой лиганд в состоянии оттянуть обычно диффузную с/-орбиту и сконцентрировать ее против себя, в результате чего возникает значительный интеграл перекрывания, а значит, и существенная энергия связи.
По относительной силе связывания атомом цинка в ферменте галогенид-ионы можно расположить в ряд 1-->ВгС1"р-, что обратно ряду по силе связывания свободного иона 2п +. Эту противоположную направленность можно интерпретировать как своего рода смягчающее влияние белка на ион цинка в противоположность сильному лиганду СН-, для которого энергия связи одинакова как со свободным так и в ферменте
При взаимодействии лиганда X с новой орбиталью (с энергией Нм) эффект связывания Дг будет меньше, чем при взаимодействии со свободным атомом металла
В отсутствие лигандов сверхспирализованные молекулы обладают большей свободной энергией, чем открытые формы двойной спирали. Доказательством тому служит тот факт, что при образовании разрыва в одиночной цепи сверхспирализация спонтанно исчезает, при этом молекула переходит в релаксированное состояние. Ббльшая величина свободной энергии в этом случае является результатом уменьшения энтропии при переходе ДНК в более компактную и более упорядоченную сверхспирализованную форму, а также увеличения энтальпии из-за появления напряжений и деформаций в структуре молекулы. Так как при < = О нативная сверхспиральная молекула находится на более высоком энергетическом уровне, чем эквивалентная ей во всем остальном открытая форма ДНК, то отсюда с необходимостью следует, что при V < свободная энергия связывания всякого лиганда, способного уменьшать число сверхвитков, должна быть меньше для сверхспира
2.1. Модельные представления, используемые для описания взаимодействия гемоглобина с кислородом
Известно, что одна молекула гемоглобина состоит из четырех мономеров, мономер имеет тем область присоединения лиганда (молекул кислорода, углекислого газа и др.) и глобин белковую часть, взаимодействующую с протонами водорода [1]. Чем может находиться в нескольких состояниях, а именно: к гему присоединен кислород; к тему присоединен углекислый газ либо он может быть свободен. Каждый гем может обратимо присоединить одну молекулу кислорода. Соответственно молекула гемоглобина может присоединить 4 молекулы кислорода.
На рисунке 12 показаны типы взаимодействий, учитываемые в модели,
где Fe ион железа, L лиганд (молекула кислорода или углекислого газа), Н протон водорода. Стрелками указаны взаимодействия, учитываемые в модели. 1 -взаимодействие лигандов с гемом, 2 кооперативное взаимодействие молекул кислорода между собой, 3 взаимодействие протонов водорода с глобином, 4 конформационное взаимодействии макромолекулы гемоглобина с протонами водорода.
Рисунок 12 – Схематичное изображение предложенной макромолекулы гемоглобина. Цифрами обозначены учитываемые взаимодействия.
Для описания модели воспользуемся базовой моделью дефектообразования в конденсированных средах, описанной в работах С. В. Булярского [2]. Данная модель вполне подходит для описания процесса взаимодействия гемоглобина с кислородом.
В соответствии с вышесказанным примем следующую модель взаимодействия гемоглобина с кислородом.
В соответствии с вышесказанным примем следующую модель взаимодействия гемоглобина с кислородом. Обозначим все возможные состояния макромолекулы гемоглобина следующей системой индексов:
{i, j, k,
} (4)Численное значение индекса i соответствует количеству присоединенных молекул кислорода, j количеству молекул углекислого газа, индекс к количеству молекул присоединение еще одного лиганда.
индексы зарядового состояния связи (донорной и акцепторной), способной принимать и отдавать водород, для аминокислоты.
2.2. Свободная энергия системы гемоглобин – лиганды
Одна из важнейших функций белков состоит в специфическом катализе химических реакций. Лигандом в этом случае служит молекула субстрата, связывание которой ферментом необходимая предпосылка химической реакции.
Ферменты способны очень сильно ускорять химические реакции значительно сильнее, чем любые искусственные катализаторы. Столь высокую эффективность можно приписать нескольким факторам. Во-первых, ферменты увеличивают локальную концентрацию молекул субстрата в каталитическом центре и удерживают соответствующие атомы в ориентации, необходимой для последующей реакции.
Но наиболее важное значение имеет тот факт, что часть энергии связывания непосредственно используется для катализа. Дело в том. что молекулы субстрата, перед тем как превратиться в продукты реакции, проходят через ряд промежуточных форм с измененной геометрией и измененным электронным распределением.
Свободная энергия всех этих промежуточных форм и особенно наименее стабильных переходных состояний существенно снижена, если молекула связана с поверхностью фермента. Обычно ферменты имеют значительно большее сродство к нестабильным переходным состояниям субстратов, чем к их стабильным формам. Используя энергию связывания, ферменты помогают субстратам принять определенное переходное состояние и таким образом значительно ускоряют одну определенную реакцию.
Свободная энергия связывания составляет —7,1 ккал/ моль, что соответствует константе сродства 10 л/моль. Присоединение лиганда перетаскивает белок К из обычной неактивной формы К в каталитически активную форму К.
И. Линейная молекула имеет очень большое число идентичных центров связывания лиганда X. Свободная энергия взаимодействия между лигандами, связанными с расположенными по соседству друг с другом центрами, равна е.
АО принимает участие в связывании с одной определенной комбинацией орбит свободных электронных пар лигандов, что приводит к накоплению примерно 0,5 единицы электронного заряда на N1 и к сравнительно небольшой энергетической стабилизации, так как обычные энергии связей бывают, по-видимому, порядка 30 ккал. Обе эти величины малы вследствие относительно большого различия в энергии между 4 и электронами свободных пар у лигандов,
Конечно, прямой доступ к иону железа для лигандов закрыт аминокислотами, особенно дистальным гистидином. Как уже отмечалось, один из атомов азота имидазольного кольца гистидина обращен к железу, а другой фактически находится на поверхности, так что этот гетероцикл может работать как своего рода люк, перекрывающий лигандную полость.
Поэтому связывание любого лиганда представляет собой сложный процесс, включающий промежуточные изменения конформации белка, например поворот гистидина Е7 вокруг его связи Са —Сз или небольшое искажение структуры спирали Е [161]. Тем не менее скорость связывания кислорода исключительно велика. Константа скорости реакции второго порядка при 20°С для различных миоглобинов находится в интервале 1,0-10 — 1,9-10 дм -моль с [определенные к настоящему времени значения свободной энергии активации для этих процессов составили в трех случаях 23,0, 23,0 и 29,3 кДж/моль (5,5, 5,5 и 7,0 ккал/моль) соответственно], а константы скорости для изолированных, но слегка модифицированных аи 3-цепей составили 5-10 — 8-10 дм моль с , тогда как для мономерного гемоглобина hironomus получено более высокое значение 3-10 дм -моль 1-с [6]. Для гемоглобинов кинетика реакции имеет сложный характер вследствие изменений четвертичной структуры, однако константы скорости и в этом случае попадают в интервал 10 — 10 дм моль с . Константы скорости отщепления кислорода составляют 10—70 с , а соответствующие энергии активации равны 80—88 кДж/моль (19—21 ккал/моль) для миоглобинов и 10— 15 с и 67—105 кДж/моль (16—25 ккал/моль) для большинства гемоглобинов (эти значения сильно зависят от pH). Библиографию по этому вопросу см. в работе [8].
Даже если гистидин существенно уменьшает величину константы скорости, которая была в отсутствие белка, наблюдаемые скорости вполне достаточны для физиологических потребностей. Мутантные гемоглобины, в которых гистидин замещен на аргинин или тирозин, обнаруживают несколько более высокие скорости, особенно в реакциях с СО [8]. Некоторые гемоглобины с очень малыми константами скорости диссоциации ( 10 с 1), которые явно не могут функционировать как переносчики кислорода, встречаются у нематод
Вывод, что вода может вступать в реакцию через 4 /-орбиту, в то время как анионы не могут сделать этого, можно
понять на основании теории Крейга о сжатии, вызываемом во внешних -орбитах электроно-притягивающими (электроотрицательными) лигандами [19]. Анионы отталкивают электроны (электроположительны), хотя в связанном виде они становятся менее электроположительными. Внешние й -орбиты не пригодны для связывания таких лигандов из-за своей большой протяженности и низкой амплитуды, вследствие чего, несмотря на полное перекрывание, интеграл перекрывания с относительно маленькими и подразделенными орбитами лигандов очень мал. Однако молекула воды, почти нейтральная в свободном состоянии, становится заряженной положительно и потому электронопритягивающей (электроотрицательной), когда она связана. Такой лиганд в состоянии оттянуть обычно диффузную с/-орбиту и сконцентрировать ее против себя, в результате чего возникает значительный интеграл перекрывания, а значит, и существенная энергия связи.
По относительной силе связывания атомом цинка в ферменте галогенид-ионы можно расположить в ряд 1-->ВгС1"р-, что обратно ряду по силе связывания свободного иона 2п +. Эту противоположную направленность можно интерпретировать как своего рода смягчающее влияние белка на ион цинка в противоположность сильному лиганду СН-, для которого энергия связи одинакова как со свободным так и в ферменте
При взаимодействии лиганда X с новой орбиталью (с энергией Нм) эффект связывания Дг будет меньше, чем при взаимодействии со свободным атомом металла
В отсутствие лигандов сверхспирализованные молекулы обладают большей свободной энергией, чем открытые формы двойной спирали. Доказательством тому служит тот факт, что при образовании разрыва в одиночной цепи сверхспирализация спонтанно исчезает, при этом молекула переходит в релаксированное состояние. Ббльшая величина свободной энергии в этом случае является результатом уменьшения энтропии при переходе ДНК в более компактную и более упорядоченную сверхспирализованную форму, а также увеличения энтальпии из-за появления напряжений и деформаций в структуре молекулы. Так как при < = О нативная сверхспиральная молекула находится на более высоком энергетическом уровне, чем эквивалентная ей во всем остальном открытая форма ДНК, то отсюда с необходимостью следует, что при V < свободная энергия связывания всякого лиганда, способного уменьшать число сверхвитков, должна быть меньше для сверхспира