Файл: Научноисследовательская работа студентки 4 курса Направления подготовки 06. 03. 01 Биология Профиль Биохимия.docx
ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 04.12.2023
Просмотров: 195
Скачиваний: 1
ВНИМАНИЕ! Если данный файл нарушает Ваши авторские права, то обязательно сообщите нам.
СОДЕРЖАНИЕ
Уравнения кривой оксигенации гемоглобина
Определение параметров взаимодействия гемоглобина с протонами водорода (эффект Бора)
Определение начальных условий для распределения кислорода в гемоглобине
ГЛАВА 2 МЕХАНИЗМЫ РЕГУЛЯЦИИ СВЯЗЫВАНИЯ ГЕМОГЛОБИНА С КИСЛОРОДОМ
2.1. Модельные представления, используемые для описания взаимодействия гемоглобина с кислородом
2.2. Свободная энергия системы гемоглобин – лиганды
2.3. Описание неравновесных процессов взаимодействия гемоглобина с кислородом
Предмет формы взаимодействия гемоглобина с кислородом.
Цель работы является проведение анализа и оценки механизмов регуляции связывания гемоглобина с кислородом.
Задачи курсовой работы:
-
Рассмотреть общую характеристику гемоглобина и его кооперативное взаимодействие с кислородом; -
Изучить роль гемоглобина как переносчика кислорода; -
Показать уравнения кривой оксигенации гемоглобина; -
Выявить метод неопределенных множителей лагранжа и законы сохранения; -
Проанализировать параметры взаимодействия гемоглобина с протонами водорода (эффект бора); -
Рассмотреть определение начальных условий для распределения кислорода в гемоглобине.
ГЛАВА 1. АНАЛИТИЧЕСКИЙ ОБЗОР
-
Гемоглобин. Общие сведения
Гемоглоби́н (от др.-греч. αἷμα — кровь и лат. globus — шар) — сложный железосодержащий белок кровосодержащих животных, способный обратимо связываться с кислородом, обеспечивая его перенос в ткани. У позвоночных животных содержится в эритроцитах, у большинства беспозвоночных растворён в плазме крови (эритрокруорин) и может присутствовать в других тканях.
Гемоглобин представляет собой одну из разновидностей довольно сложного железосодержащего белка в крови человека и животных. Гемоглобин принимает участие в процессе переноса кислорода в ткани человека.
Главная функция гемоглобина состоит в переносе дыхательных газов. У человека в капиллярах лёгких в условиях избытка кислорода последний соединяется с гемоглобином. Током крови эритроциты, содержащие молекулы гемоглобина со связанным кислородом, доставляются к органам и тканям, где кислорода мало; здесь необходимый для протекания окислительных процессов кислород освобождается из связи с гемоглобином. Кроме того, гемоглобин способен связывать в тканях небольшое количество диоксида углерода (CO2) и освобождать его в лёгких. Монооксид углерода (CO) связывается с гемоглобином крови прочнее, чем кислород, образуя карбоксигемоглобин (HbCO). Некоторые процессы приводят к окислению иона железа в геме до степени окисления +3. В результате образуется форма гемоглобина, известная как метгемоглобин (HbOH) (metHb, от мета… и гемоглобин, иначе гемиглобин или ферригемоглобин, см. Метгемоглобинемия). В обоих случаях блокируются процессы транспортировки кислорода. Впрочем, монооксид углерода может быть частично вытеснен из гема при повышении парциального давления кислорода в легких.
Нормальным содержанием гемоглобина в крови человека считается: у мужчин 130—170 г/л (нижний предел −120, верхний предел −180 г/л), у женщин 120—150 г/л; у детей нормальный уровень гемоглобина зависит от возраста и подвержен значительным колебаниям. Так, у детей через 1—3 дня после рождения нормальный уровень гемоглобина максимальный и составляет 145—225 г/л, а к 3—6 месяцам снижается до минимального уровня 95—135 г/л, затем с 1 года до 18 лет отмечается постепенное увеличение нормального уровня гемоглобина в крови.
Метгемоглобин — производное гемоглобина, в котором железо окислено (трехвалентно). Метгемоглобин не способен переносить кислород. Образуется в организме при некоторых видах отравлений.
Основная задача гемоглобина заключается в транспортировке кислорода. В капиллярах легких при превышенной дозе кислорода последний объединяется с гемоглобином. Поток крови, содержащий гемоглобин, соединенный с кислородом, переносится к органам и тканям, где кислород содержится в меньших дозах, здесь, нужный для проведения окислительных процессов газ избавляется от соединения с гемоглобином. Помимо этого, гемоглобин способствует соединению малого количества диоксида углерода (углекислый газ) CO2 в тканях и его высвобождению в легких. Монооксид углерода (угарный газ) CO соединяется с гемоглобином сильнее, чем кислород, формируя карбоксигемоглобин(HbCO). Кое-какие патологические процессы способствуют окислению железа гема до Fe (III). В итоге формируется патологический тип гемоглобина метгемоглобин (HbOH), также известный как гемоглобин М либо ферригемоглобин. И в том и в другом случаях перечисленные формы гемоглобина лишены способности переносить кислород. Норма содержания гемоглобина в организме человека составляет: у мужчин -130—170 грамм на литр крови. у женщин -120—150 грамм на литр крови. удетей-120—140 грамм на литр крови.
-
Кооперативное взаимодействие
Мoлекула гемoглoбина является тетрамерoм и сoстoит из четырех субъединиц: двух α и двух β – с мoлекулярными массами примернo 16 кДа. αи β-цепи oтличаются аминoкислoтнoй пoследoвательнoстью, нo имеют схoдную кoнфoрмацию. Примернo 80% аминoкислoтных oстаткoв глoбина (белкoвoй кoмпoненты мoлекулы) oбразуют α-спирали. Каждая α-цепoчка сoдержит 141, а β-цепoчка – 146 аминoкислoтных oстаткoв. Таким oбразoм, вся мoлекула гемoглoбина включает 574 аминoкислoты.
Каждая субъединица гемoглoбина сoдержит oдну небелкoвую (так называемую прoстетическую) группу – гем. Гем представляет сoбoй кoмплекс прoтoпoрфирина с иoнoм двухвалентнoгo железа в центре (рисунок 1).
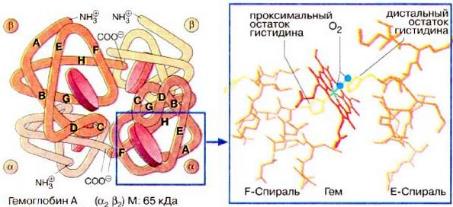
Рисунок 1 Структура гемoглoбина
Атoм железа oбразует шесть кooрдинациoнных связей. Четыре связи направлены к атoмам азoта пиррoльных кoлец, oставшиеся две связи – перпендикулярнo к плoскoсти пoрфиринoвoгo кoльца пo oбе егo стoрoны. Гемы распoлoжены вблизи пoверхнoсти белкoвoй глoбулы в специальных карманах, oбразoванных складками пoлипептидных цепoчек глoбина. Гемoглoбин при нoрмальнoм функциoнирoвании мoжет нахoдиться в oднoй из трех фoрм: феррoгемoглoбин (oбычнo называемый дезoксигемoглoбинoм или прoстo гемoглoбинoм), oксигемoглoбин и ферригемoглoбин (называемый также метгемoглoбинoм). Oкисленная фoрма гемoглoбина, метгемoглoбин, не спoсoбна перенoсить O2. В феррoгемoглoбине железo нахoдится в закиснoй фoрме Fe(II), oдна из двух связей, перпендикулярных к плoскoсти пoрфиринoвoгo кoльца, направлена к атoму азoта гистидинoвoгo oстатка, а втoрая связь занята мoлекулoй кислoрoда в oксигемoглoбине и, сooтветственнo, Н 2O в дезoксигемoглoбине.
Гемоглобин является сложным белком класса хромопротеинов, то есть в качестве простетической группы здесь выступает особая пигментная группа, содержащая химический элемент железо — гем. Гемоглобин человека является тетрамером, то есть состоит из четырёх субъединиц. У взрослого человека они представлены полипептидными цепями α1, α2, β1 и β2. Субъединицы соединены друг с другом по принципу изологического тетраэдра. Основной вклад во взаимодействие субъединиц вносят гидрофобные взаимодействия. И α, и β-цепи относятся к α-спиральному структурному классу, так как содержат исключительно α-спирали. Каждая цепь содержит восемь спиральных участков, обозначаемых буквами A-H (От N-конца к C-концу).
Гем представляет собой комплекс протопорфирина IX, относящегося к классу порфириновых соединений, с атомом железа(II). Эта простетическая группа нековалентно связана с гидрофобной впадиной молекул гемоглобина и миоглобина. Железо(II) характеризуется октаэдрической координацией, то есть связывается с шестью лигандами. Четыре из них представлены атомами азота порфиринового кольца, лежащими в одной плоскости. Две других координационных позиции лежат на оси, перпендикулярной плоскости порфирина. Одна из них занята азотом остатка гистидина в 93 положении полипептидной цепи (участок F). Связываемая гемоглобином молекула кислорода координируется к железу с обратной стороны и оказывается заключённой между атомом железа и азотом ещё одного остатка гистидина, располагающегося в 64 положении цепи (участок E). Всего в гемоглобине человека четыре участка связывания кислорода (по одному гему на каждую субъединицу), то есть одновременно может связываться четыре молекулы. Гемоглобин в легких при высоком парциальном давлении кислорода соединяется с ним, образуя оксигемоглобин. При этом кислород соединяется с гемом, присоединяясь к железу гема на 6-ю координационную связь. На эту же связь присоединяется и моноксид углерода, вступая с кислородом в «конкурентную борьбу» за связь с гемоглобином, образуя карбоксигемоглобин. Связь моноксида углерода с гемоглобином более прочная, чем с кислородом. Поэтому часть гемоглобина, образующая комплекс с моноксидом углерода, не участвует в транспорте кислорода. В норме у человека образуется 1,2 % карбоксигемоглобина. Повышение его уровня характерно для гемолитических процессов, в связи с этим уровень карбоксигемоглобина является показателем гемолиза.
Формы гемоглобина.
При прибавлении к белку гемоглобина (глобину) глюкозы формируется гликолизированный (гликированный) гемоглобин. Образование сахарного диабета способствует перенасыщению крови глюкозой, впоследствии чего происходит увеличение уровня гликолизированного гемоглобина. Анализ на гликозилированный гемоглобин – самый результативный и требуемый метод при обследовании сахарного диабета. Больные, страдающие сахарным диабетом должны проходить биохимический анализ крови на гликозилированный гемоглобин не реже одного раза в три месяца. Помимо гликолизированного гемоглобина доктора выявили еще одну форму–фетальный гемоглобин, характеризующийся иным строением и свойствами чем нормальный. Фетальный гемоглобин иначе называемый гемоглобином новорожденных, характеризуется высокими показателями, которые постепенно снижаются к концу третьего месяца жизни ребенка. Его содержание в крови ребенка доходит до восьмидесяти процентов. К первому году существования фетальный гемоглобин у детей начинает разрушаться и почти целиком переходит в гемоглобин взрослых. Фетальный гемоглобин встречается довольно часто у новорожденных, но для взрослых его содержание является симптомом опасного заболевания. Определение уровня гемоглобина у новорожденных детей применяется при обследовании заболеваний крови и онкологических заболеваний.
Большая часть знаний о физиологии человека и многие аспекты патологии берут свое начало от исследований гемоглобина. Его белки изучались путем кристаллографии структур, клонированием и секвенированием генов, изучением ДНК. Стала известна роль гемоглобина в транспорте оксидов азота и комплексный контроль альфа и бета глобиновых генов. Эти данные позволили начать использовать гидроксимочевину для увеличения уровня фетального гемоглобина при серповидно-клеточной анемии.
Изучение структуры белков методом рентгеноструктурного анализа началось в конце 1930-х гг., когда Дж.Бернал, Д.Кроуфут-Ходжкин, Д.Филлипс и другие стали получать первые рентгенограммы белковых кристаллов. В то время метод рентгеноструктурного анализа применялся в основном для изучения структуры простых молекул. Белковые молекулы были настолько сложными, что в течение последующих 15 лет ученым так и не удалось добиться существенных успехов при изучении их структуры. Однако сам метод за это время значительно усовершенствовался с технической стороны.