ВУЗ: Не указан
Категория: Не указан
Дисциплина: Не указана
Добавлен: 15.09.2020
Просмотров: 5898
Скачиваний: 284

нями деревьев не только более глубоких слоев почвы, но в пони-
женных местах и самого верхнего слоя, в результате чего создают-
ся «мертвые» для корней деревьев зоны, занимающие большую
или меньшую часть всей поверхности в зависимости от общего
режима переувлажнения данного участка.
2.6.
Биогеоценозы подзоны хвойно-
широколиственных лесов
Уникальными для центра России являются ландшафты так
называемого
ополья —
наиболее древнеосвоенные земли с пло-
дородными почвами в бассейне некоторых озер, на плоских озер-
но-ледниковых и озерно-водноледниковых равнинах. Таковы тер-
ритории вокруг озер Неро, Плещеево, некоторые равнинные
плато Владимирской, Ивановской, Тверской и Московской об-
ластей. В зональном плане ополья занимают пограничную тер-
риторию, представляя собой самую северную часть ареала хвой-
но-широколиственных лесов в данной провинции Русской рав-
нины. Ландшафты ополий — это волнистые слабо и среднедре-
нированные возвышенные равнины с преобладанием серых лес-
ных легкосуглинистых почв на покровных лессовидных суглин-
ках.
В силу давнего освоения и почти повсеместного сведения лесов
(еще на рубеже I — II тысячелетий) реконструировать ландшафт-
ную структур подтаежной зоны можно лишь с известной долей
условности. На покровных суглинках вторичных моренных и вод-
но-ледниковых равнин водораздельные местоположения во всех
ярусах были заняты
елово-широколиственными лесами,
обильно
насыщенными представителями неморальной флоры. Основным
эдификаторами (строителями биоценозов) этих лесов были ель
европейская и липа мелколистная.
В условиях хорошо выраженного рельефа конечно-моренных
гряд на серых лесных почвах когда-то возникли леса богатого ден-
дрологического состава, где помимо ели европейской и липы мел-
колистной большое участие в первом ярусе принимает дуб череш-
чатый, а во втором — ильм (вяз) и клен, а иногда (на карбонат-
ной морене) и ясень.
На более плоских и волнистых пространствах вторичных мо-
ренных равнин с периодическим сезонным переувлажнением вер-
хних горизонтов почвы усиливаются процессы оподзоливания, и
здесь, на подзолистых и светло-серых почвах, существовали леса
из липы и ели с единичными экземплярами дуба в первом ярусе.
На периферии моренных и водно-ледниковых водоразделов в ус-
ловиях еще более затрудненного дренажа развивались еловые леса
с липой во втором ярусе или в подлеске (рис. 2.18).

Распространившиеся теперь чрезвычайно широко
осинники
первоначально занимали лишь наиболее богатые местообитания
переувлажненных водораздельных депрессий и котловин. Нако-
нец, замкнутые понижения равнин, где подолгу задерживаются
верховодка, были заняты коренными лесами из березы пуши-
стой.
В пределах широких зандровых полей господствовали чисто хвой-
ные насаждения южно-таежного типа с участием ели и сосны и
совершенным отсутствием неморальной флоры — так проявля-
лась своеобразная «экспансия тайги» на территорию более южной
зоны.
На древнеаллювиальных отложениях высоких речных террас
также были распространены боровые урочища. Достопримечатель-
ностью территории являются сосново-широколиственные леса,
которые, правда, иногда относят к производным, т.е. изменен-
ным человеком, урочищам.
Сосново-широколиственные леса
занимают в настоящее время
наиболее бедные маломощные супеси, подстилаемые мореной.
Здесь в травяном покрове преобладают отчетливо бореальные
формы, но во втором ярусе древостоя участвуют липа и дуб.
В подлеске — бересклет, жимолость, рябина. На более богатых
местообитаниях распространены сосняки с участием ели и широ-
колиственных пород и с господством неморальных видов в назем-
ном покрове.
По проточными мокрым низинам вблизи ручьев так же, как и
теперь, были широко распространены черноольховые леса. При-
чем при хорошей проточности формировались высокобонитетные
черноольшаники с богатым наземным покровом с участием дуба
и вяза в первом ярусе. До нашего времени уцелели лишь низкобо-
нитетные черноольховые леса с примесью березы и дерновинны-
ми осоками в наземном покрове.
В верховьях рек по берегам водотоков росли черноольховые леса.
В низовьях на долгопоемных участках — осокоревые и ивовые. На
центральных поймах — вязовые. Все они к нашему времени совер-
шенно исчезли — остались заросли ивняка и сероольшаники. Не
дошли до нас и дубовые рощи высоких пойм.
Позиции любых широколиственных пород на всем севере ев-
ропейской территории России (ЕТР) достаточно непрочны. Су-
ществуя в экстремальных условиях, деревья подвергаются разли-
чным воздействиям, среди которых наиболее жесткими можно
считать чередование глубоких влажных оттепелей с мокрым сне-
гом и дождем с резкими (как правило, ночными) похолодания-
ми. Набухшая от воды кора при заморозках лопается и дает так
называемые морозобойные трещины длиной в несколько метров
по всей комлевой части ствола. В летнее время морозобои играют
роль проходов для насекомых, в них же поселяются грибы.
Qfl
Знаковым деревом широколиственного древостоя традицион-
но считается
дуб черешчатып.
Существование чистых коренных
водораздельных дубовых рощ в пределах подзоны проблематично
и оспаривается многими специалистами. Большая часть сохранив-
шихся на водоразделах дубовых рощ являются саженными, даже в
том случае, если сведения об их культурном происхождении ни-
кому не известны. Более вероятным считается естественное про-
исхождение
пойменных дубрав,
в частности в Верхневолжье и на
всем северо-западе ЕТР сохранились фрагменты такого рода до-
линно-речных биогеоценозов. Древесина дуба часто встречается в
разрезах погребенных пойменных почв, переживших стадию на-
илконакопления. «Парковые» дубовые рощи сохранились кое-где
и на гривах самой Волги, хотя естественность их происхождения
остается дискуссионной. С одной стороны, естественное проис-
хождение дубовых молодняков (в возрасте 20 — 30 лет) не вызы-
вает никакого сомнения, с другой — проведенные историко-гео-
графические исследования свидетельствуют, что практически все
старые (более чем 100-летние) массивы такого рода были окру-
жены заботой и уходом.
С.Ф.Курнаев — автор классической работы по хвойно-широ-
колиственным лесам Русской равнины [26] — полагал, что нет ни
одного описания чистой водораздельной дубравы из центральных
областей, которая имела бы свойства бесспорного самобытного
ценоза. Преобладание дуба в первом ярусе хвойно-широколиствен-
ных древостоев он объяснял повсеместным выбиранием другой
широколиственой породы —
липы мелколистной
в ходе прииско-
вых рубок, длившихся столетиями, поскольку известно, что взрос-
лые экземпляры липы шли на разнообразную «столярку», а под-
рост уничтожался в результате обдирания коры «на лыко». Раз-
личными авторами также высказывалось мнение о дубе как о за-
носной для лесов Восточной Европы породе. Продвижение дуба
на север связывают с обезлесением, созданием крупных массивов
сельскохозяйственных угодий, подсек в том числе, и обогащени-
ем почвы.
Чистые липовые древостой встречаются ныне крайне редко, а
все исследованные нами старовозрастные рощи оказывались ос-
татками бывших дворянских усадеб или старых сельских парков,
хотя в отдельных случаях заросший липовый парк с активным
возобновлением молодых деревьев, влажным крупнотравьем и
практически сомкнутым пологом мощных крон может выглядеть
как естественное насаждение. Однако опытный глаз найдет отли-
чия между культурными насаждениями и «дикой» рощей: в пар-
ках боковые ветви лип в нижней части ствола обычно обрезали до
их естественного выпадения, ибо в последнем случае на месте
отпавшей ветви возникали глубокие дупла, которые могли приве-
сти к заболеванию и ранней гибели дерева.
91

В отличие от липы и дуба
клен платановидный
всегда остается
деревом второго яруса и практически не образует собственных
насаждений. Способность клена к восстановлению на вырубках (и
вообще при осветлении полога леса) поразительна — встречают-
ся участки леса, где одновозрастный кленовый подрост достигает
густоты нескольких экземпляров на квадратный метр. В. И.Турма-
нина [48] считает, что экспансия клена происходит все последнее
столетие.
Этот же автор полагает, что для вяза XX в. стал роковым, оба
схожих по экологии вида этого рода: вяз гладкий и вяз шершавый —
скорее всего проникли в естественные древостой из парковых лан-
дшафтов, однако в последние десятилетия вязы гибнут вследствие
распространения грибкового заболевания — так называемой гол-
ландской болезни вяза.
Ясень
также весьма активно заполняет окна во втором ярусе
древостоя хвойно-широколиственных лесов. Обильная поросль
ясеня близи материнских деревьев объясняется корнеотпрыско-
вым возобновлением этой породы.
Еще одним характерным признаком хвойно-широколиствен-
ных лесов является присутствие так называемой
неморальной сви-
ты,
в которую входят кустарники и травы — спутники широко-
лиственных древостоев. Из кустарников наиболее типична лещина
(орешник), нередко образующая заросли высотой до 5 м и даже
формирующая третий ярус в елово-широколиственных лесах. Ви-
дов трав, относимых к неморалам, довольно много, большинство
из них, по мнению В. И.Турманиновой, когда-то входили в свиту
дуба и липы, но в дальнейшем при деградации древостоев после-
дних смогли пережить неблагоприятные условия и оторвались от
своих средообразователей. К таким растениями относится широко
распространенный практически на всем севере ЕТР ландыш. Точ-
но также ветреница дубравная, перелеска, чина весенняя, звезд-
чатки, злаки перловник и бор развесистый сегодня прекрасно
чувствуют себя и в типичных южно-таежных травяно-кисличных
ельниках, без всякой примеси широколиственных пород
(ельники
неморальные).
К дубравным видам, и сейчас сохраняющим прочную связь с
широколиственным породами, относятся копытень, пролесник
многолетний, подмаренник душистый, лютик кашубский и не-
которые другие.
Ландшафты зоны хвойно-широколиственных лесов чрезвычайно
динамичны, что обусловлено особыми свойствами биоты. Древе-
сные породы, составляющие биоценозы хвойно-широколиствен-
ных лесов, обладают различной долговечностью и темпами роста.
Наиболее долговечна липа, которая при благоприятных условиях
доживает до 250 — 280 лет. Еловые древостой могут находиться в
здоровом состояния (без признаков распада) вплоть до 170 — 200
92
лет. Дуб менее долговечен, так как находится у северных пределов
своего ареала — деревья 200-летнего возраста, как правило, сухо-
вершинные и имеют гнилую сердцевину.
Породы второго яруса: вяз, клен, ильм — редко достигают воз-
раста 120 лет, причем все старые деревья бывают поражены моро-
зобойными трещинами. Незначительное число деревьев погибает,
так и не достигнув зрелого возраста. Наименее долговечны береза
(100 лет), осина (80 — 90 лет), ольха (70 лет).
Понятно, что совместное существование столь разных видов
в одном сообществе возможно только благодаря куртинному их
размещению: округлые куртины господствующих пород мозаич-
но чередуются в ландшафте, все прочие породы: дуб, вяз, береза,
рябина — разбросаны одиночными деревьями в просветах (ок-
нах) между куртинами. Куртинность леса является коренным
свойством биоты хвойно-широколиственных ландшафтов и вы-
ражается наиболее четко на участках давно не подвергавшихся
вырубкам.
Контрольные вопросы и задания
1. В табл. 2.2 дан ключ к определению урочищ сосновых лесов на пес
чаных и супесчаных почвах водно-ледниковых, древнеаллювиальных рав
нин и надпойменных террас для центра Русской равнины. Проанализи
руйте логику использования ключа и сопоставьте описания биогеоцено
зов с приведенными ниже названиями урочищ: сосняк лишайниковый,
сосняк бруснично-зеленомошный, сосняк молиниевый, сосняк-чернич
ник, сосняк багульниково-сфагновый, сосняк пушицево-сфагновый,
сосняк-долгомошник, сосняк орляковый, сосняк вересковый, сосняк
овсяницевый.Заполните ячейки.
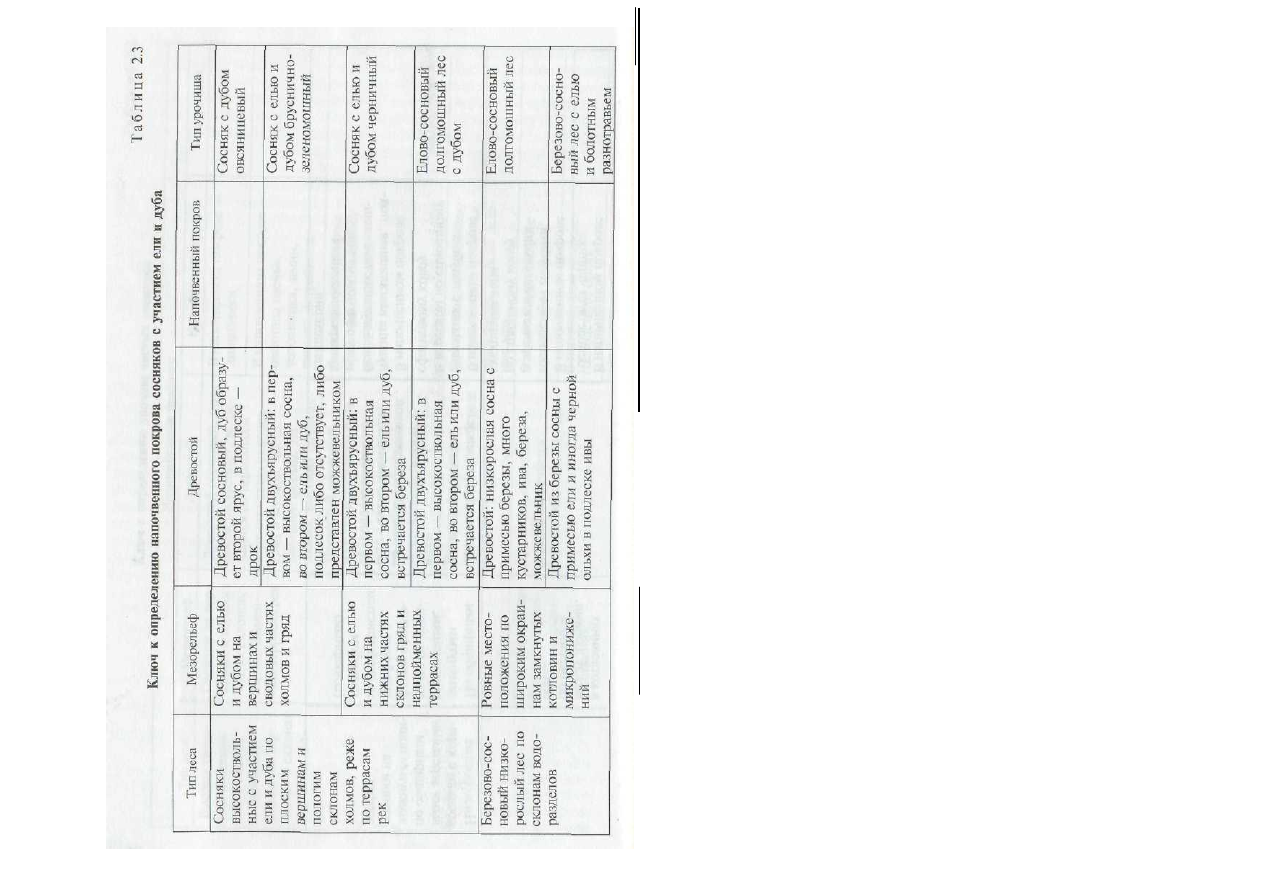
2. Заполните ячейки табл. 2.3, относящиеся к характеристике напо
чвенного покрова биогеоценозов в ключе для определения урочищ сме
шанных сосновых лесов центра Русской равнины.
Характеристики напочвенного покрова:
• брусника по сплошному покрову мхов, дрок, черника, орляк, вей-
ник;
•
моховой покров из мхов-долгомошников с пятнами сфагнума, брус
ника, _ голубика;
•
травяной покров густой с господством тростника, осок, часто —
папортники, хвощ, вербейники;
• травостой разреженный: овсяница овечья, кошачья лапка, иногда —
лишайники;
• сплошной покров черники, молиния, брусника, моховой покров из
зеленых мхов;
• в травостое осоки, ситник, голубика, подбел, вахта, сабельник.
3. Используя характеристики экологических рядов, составьте само-
стоятельно ключ для определения урочищ еловых лесов подзоны южной
тайги для центра Русской равнины.
93

Глава 3
БОЛОТНЫЕ ЛАНДШАФТЫ И ФАКТОРАЛЬНО-
ЭКОЛОГИЧЕСКИЕ РЯДЫ БОЛОТНЫХ БИОГЕОЦЕНОЗОВ
3.1.
Генезис и классификация болотных
ландшафтов
Болота и заболоченные леса — настолько характерная черта
ландшафтного облика центра и севера европейской России, что
мы не можем не уделить им особого внимания. Генезис болот тес-
но связан с процессами дегляциации территории со времени пос-
леднего ледникового покрова. Разнообразие форм унаследованно-
го рельефа, различие в степени дренированности территории, а
также многообразие форм застоя атмосферной влаги и «раскры-
тия» грунтовых вод определили разнообразие условий болотооб-
разования. В этом смысле каждое болото уникально — по рельефу,
условиям грунтового и атмосферного питания, почвам и развива-
ющимся биогеоценозам.
Трудности в типологии болот и их классифицировании также
связаны с чрезвычайным разнообразием условий болотообразо-
вания.
Принятое деление болот на верховые (заросшие и запол-
ненные торфом котловины послеледниковых озер) и низинные
(заторфованные депрессии в местах выхода грунтовых вод) под-
ходит лишь для своего рода «крайних» членов ряда, огромная же
масса реальных болотных массивов лежит где-то посередине меж-
ду этими крайними типами, имеет смешанное питание, разли-
чный мезо- и микрорельеф и разнообразный набор растительных
ассоциаций.
Специфика и типология растительных ассоциаций болот за-
висит от множества факторов, как абиогенных (условия пита-
ния, рельеф, подстилающие породы), так и биогенных. Ланд-
шафтный облик болот именно в силу этих причин неповторим и
своеобразен, кроме того, многие болота находятся на разных
стадиях биоценотических смен (сукцессии). На примере болот
мы сталкиваемся с ситуацией, когда мезорельеф (геотопическая,
пространственная координата) перестает быть единственным
ведущим фактором дифференциации ландшафтной оболочки,
поскольку важное значение приобретают как иные факторы (тип
и характер увлажнения, состав подтекающих грунтовых вод), так
97